Myocardium and Heart
A merry heart doeth good like a medicine.
—Proverbs XVII:22
Objectives for This Exercise
Exercise on Blood Vessels
Slides For This Exercise: 616, 5-20, 582, 582A, 525, 102, 96, 1-75, MCO0033
THERE IS A PRE-RECORDED LECTURE FOR THIS EXERCISE
Please go to the listing page for these to download and view it
Strictly speaking, the term "cardiovascular system" isn't synonymous with
"circulatory system," since the latter technically includes the
lymphatic, as well as the blood, circulation. Of course, the two terms are
often used interchangeably, and you should be alert for this when consulting
the literature. In this exercise we'll be concerned strictly with the blood circulatory
apparatus.
The cardiovascular system can be anatomically subdivided into the heart
and the blood vessels, and the latter category is further subdivided
into different vessel types.
Arteries are blood vessels which conduct blood away from the heart; veins
are blood vessels which conduct blood towards the heart. These two major
categories are broken down further, and there are subcategories of arteries and
veins based on size, construction of the vessel wall, etc. However, the
direction of blood flow is the criterion which separates arteries from veins,
and which defines a vessel as being in one or the other major class. You may
have heard the hoary old chestnut that "Arteries carry oxygenated blood,
and veins carry deoxygenated blood." THIS IS NOT TRUE. Some
arteries carry deoxygenated, and some veins carry oxygenated blood. The
direction of flow is the only criterion for classification.
We'll look at blood vessels later in another exercise,
but for now, let's concentrate on the heart.
The Heart
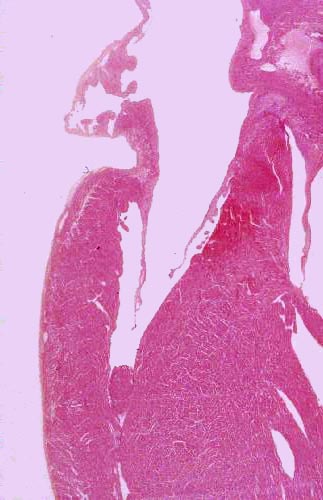
 Begin with slide 616. This is an entire
heart from some small animal (most likely a rat) which has been sectioned from
the apex to the cranial end of the atria. After orienting yourself, examine it at very low power. You will easily be able to make out at least
two, and probably three of the chambers; if you have a favorable section, you
may be able to see all four. The atria and ventricles are separated from each
other. Running from the apex to the cranial end you will see the septum
dividing the right and left sides. There should be at least one of the heart
valves in your section, connecting the atria and ventricles. External to the
heart proper, you should have a cross section of one of the great vessels (the
pulmonary artery) and some adipose connective tissue as well.
Begin with slide 616. This is an entire
heart from some small animal (most likely a rat) which has been sectioned from
the apex to the cranial end of the atria. After orienting yourself, examine it at very low power. You will easily be able to make out at least
two, and probably three of the chambers; if you have a favorable section, you
may be able to see all four. The atria and ventricles are separated from each
other. Running from the apex to the cranial end you will see the septum
dividing the right and left sides. There should be at least one of the heart
valves in your section, connecting the atria and ventricles. External to the
heart proper, you should have a cross section of one of the great vessels (the
pulmonary artery) and some adipose connective tissue as well.
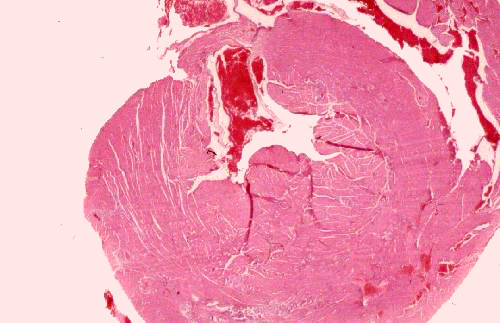 Turn now to slide 66. This is the heart of yet another unfortunate rodent, sectioned transversely, below the level of the atrioventricular
junctions. This slide graphically illustrates
the thickness of the wall of the left ventricle and ventricular septum. The cross section at right shows a "slug" of blood frozen in its passage from the left atrium to the left ventricle, and the left A-V valve is open. On this slide you should be able to make out
most of the features identified in the previous slide, except those which are
out of the plane of the section.
Turn now to slide 66. This is the heart of yet another unfortunate rodent, sectioned transversely, below the level of the atrioventricular
junctions. This slide graphically illustrates
the thickness of the wall of the left ventricle and ventricular septum. The cross section at right shows a "slug" of blood frozen in its passage from the left atrium to the left ventricle, and the left A-V valve is open. On this slide you should be able to make out
most of the features identified in the previous slide, except those which are
out of the plane of the section.
While both atria and ventricles are composed of the same type of specialized muscle cells, those of the atria tend to be less numerous, thinner, and more elongated than those of the ventricles. The ventricles of the heart are more stoutly constructed than the atria because they have more work to do. All the atria do is pass blood to the ventricles below: the right ventricle sends blood to the lungs against the resistance of the pulmonary circulation, and the the left ventricle has to deal with the entire system circulation's resistance to flow. While the resistance of the lung capillaries to blood flow is considerable, that of the entire peripheral circulation is much higher.
Myocardium
The heart is a large mass of muscle. Cardiac muscle is a variant form of striated muscle, with distinct differences from the skeletal form; and some unique structures that make it work properly, day in and day out, until death. The term "myocardium," (from Greek, myos = muscle + kardio = heart) is specifically the mass of muscle that comprises the bulk of the organ.
On slides 616 and 66 at low power, you can easily observe the pattern of the muscle
cells of the myocardium. Myocardial cells (cardiac myocytes) are much smaller than the
myofibers of skeletal muscle, and they don't form long cylindrical structures
the way myofibers do. While skeletal myofibers can in some cases be meters
in length, it's a rare cardiac myocyte that exceeds 100 µm in length, and
perhaps 30-50 µm wide. The shape of the myocyte is pretty irregular, with
stumpy projections coming off of it where it contacts other cells. Myocytes also differ from skeletal myofibers in that cardiac myocytes are mononuclear and not syncytial. One nucleus per myocyte is the rule, not hundreds as in a skeletal myofiber, and that one nucleus is centrally located.
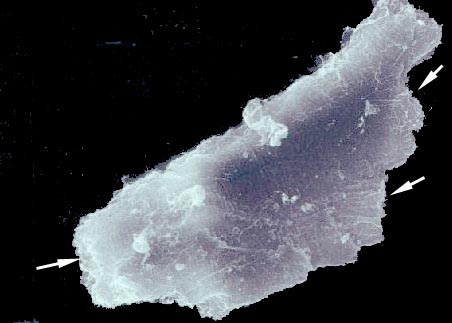 Here's scanning electron microscope image of a single cardiac myocyte, which has been mechanically separated from the mass of the myocardium. This beautiful image (at about 5000x) shows the irregular shape of the myocyte quite well, and the points at which this cell is in contact with others via the intercalated discs are indicated by arrows. Also visible as wispy strands of material on the surface are fine collagen fibrils of the intercellular collagen network. Just as in skeletal muscle, the ICN transmits force throughout the mass, and the hierarchical arrangement of endomysium-perimysium-epimysium applies.
Here's scanning electron microscope image of a single cardiac myocyte, which has been mechanically separated from the mass of the myocardium. This beautiful image (at about 5000x) shows the irregular shape of the myocyte quite well, and the points at which this cell is in contact with others via the intercalated discs are indicated by arrows. Also visible as wispy strands of material on the surface are fine collagen fibrils of the intercellular collagen network. Just as in skeletal muscle, the ICN transmits force throughout the mass, and the hierarchical arrangement of endomysium-perimysium-epimysium applies.
Note that the surface of the myocyte appears to be ridged or grooved. These ridges are the sites of Z-lines. They mark the ends of the sarcomeres of this cell's contractile apparatus. The sarcomeres are slightly contracted in this specimen and so they appear as ridges on the surface. The T-tubules aren't visible.
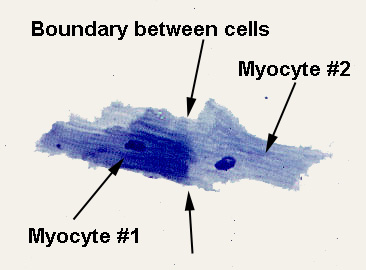 In the image at right, a whole mount stained with toluidine blue at about 1000x, two myocytes have come through the disaggregation procedure but remain connected to each other: one is more heavily stained than its partner, so that the boundary where the two are joined (an intercalated disk, see below) is clear. As is obvious in this image, each cell has a single centrally located nucleus. Unlike skeletal muscle, neither myocardium nor the cells of which it's composed are syncytial. The cells are physically isolated from each other, a fact which was in dispute before the electron microscope settled the matter once and for all about 1950. (Nevertheless, thanks to the Law Of Persistence Of Erroneous Information, you'll still find the statement in texts that "the heart is functionally syncytial," something that sets my teeth on edge.) Though there is undoubtedly constant communication and coordination between heart muscle cells, they are structurally distinct entities, and the term "syncytium" is inappropriate in this context.
In the image at right, a whole mount stained with toluidine blue at about 1000x, two myocytes have come through the disaggregation procedure but remain connected to each other: one is more heavily stained than its partner, so that the boundary where the two are joined (an intercalated disk, see below) is clear. As is obvious in this image, each cell has a single centrally located nucleus. Unlike skeletal muscle, neither myocardium nor the cells of which it's composed are syncytial. The cells are physically isolated from each other, a fact which was in dispute before the electron microscope settled the matter once and for all about 1950. (Nevertheless, thanks to the Law Of Persistence Of Erroneous Information, you'll still find the statement in texts that "the heart is functionally syncytial," something that sets my teeth on edge.) Though there is undoubtedly constant communication and coordination between heart muscle cells, they are structurally distinct entities, and the term "syncytium" is inappropriate in this context.
Thanks to the difference of cellular architecture, the histological appearance of myocardium is very different from that of skeletal muscle. Rather than forming neat bundles of parallel fibers with well-defined striations, myocardium forms an anastomosing network with a sort of "spongy" appearance. The contacts between myocytes are such that a branching network with blood vessels in the spaces between them is the result.
You will see some areas where the myocardial cells are oriented end-to-end in
long rows parallel to the plane of the section (i.e., they are cut
longitudinally) and others where they run at right angles (and thus are cut
transversely). There will also be areas where the orientation with respect to
the plane of the section is less well defined; these are oblique sections. The
myocardial muscle bundles are oriented in such a way as to make most efficient
use of the force of contraction.
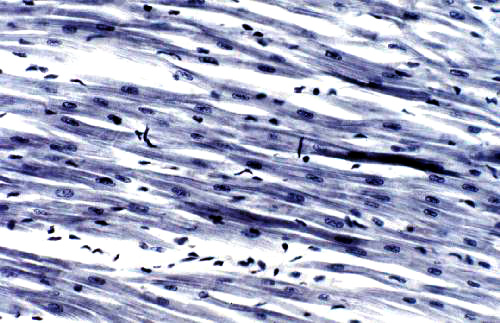
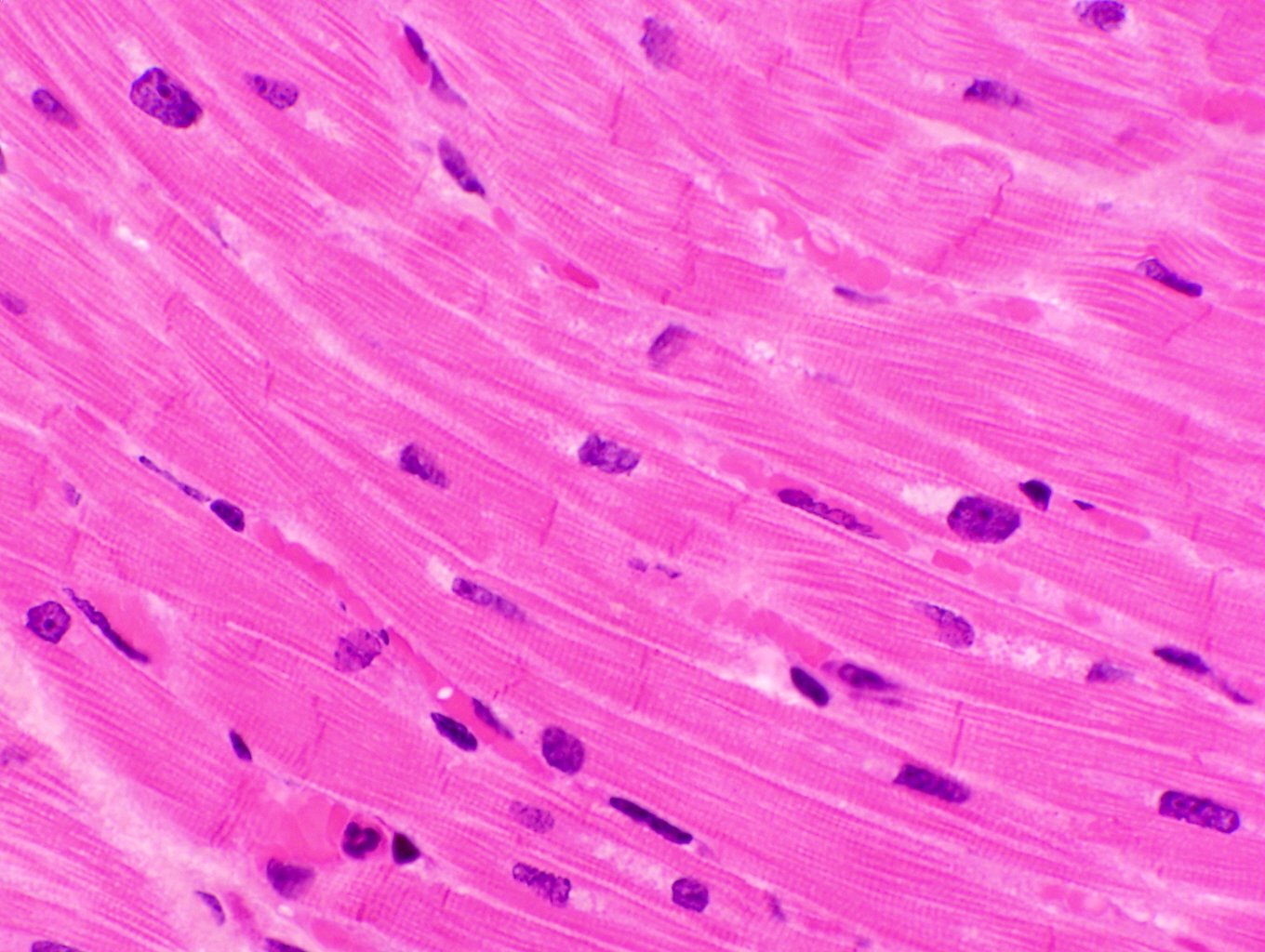 These two images give a pretty good idea of the appearance of myocardium in the light microscope. The one at left is stained with toluidine blue at about 200x; the one at right with H&E at roughly 400x.
These two images give a pretty good idea of the appearance of myocardium in the light microscope. The one at left is stained with toluidine blue at about 200x; the one at right with H&E at roughly 400x.
Myocardium has a "stringy" look compared to skeletal muscle. While skeletal muscle cells are very large and lie next to each other in parallel bundles, the smaller cardiac muscle cells are butted together at their ends, and irregularly shaped, numerous blood vessels between them. The effect is to create an anastomosing network of fibers rather than a solid phalanx of muscle fascicles. The single nucleus of each myocardial cell is quite clearly visible in these images, especially on the right.


This drawing will clarify the architecture of the tissue. Note the branching, and compare it to the actual specimens shown in the longitudinal view. At lower right in the drawing, the cells are shown in cross section. Compare the drawing to the actual specimen shown at right, and to the sections of smooth muscle in Exercise 10: at first glance this field could be confused with smooth muscle, but they can be told apart fairly easily. An important difference between the appearance of this tissue compared to smooth muscle is that in cardiac muscle almost all the cell profiles are approximately the same size, which isn't true of smooth muscle. Furthermore, most of the cell profiles will have a nucleus inside. Cardiac myocytes are so short and the nucleus takes up such a proportionally greater percentage of their length (compared to smooth muscle cells) that the chances of intersecting a nucleus with the plane of the section is pretty good. The individual cells are clearly defined by their CT envelope (the endomysium) and most of them show a nuclear profile.
The smaller, denser nuclei outside the cells are fibroblasts that make and maintain the intercellular collagen network. There's a blood vessel crossing the field about one quarter of the way down from the top edge. Cardiac muscle is even better served by the circulatory system than skeletal muscle is. Unlike skeletal muscle, cardiac muscle can't incur an "oxygen debt," because it doesn't have the lactic acid pathway to generate ATP when oxygen levels are low. Consequently it's quite prone to anoxia, and the capillary beds are extensive to minimize the possibility of it happening.
Intercalated Disks
The most prominent feature of myocardium, and the one that's absolutely diagnostic for it, is the intercalated disk. The ID is a specialized cell-to-cell adhesion/communications site. It demarcates the beginning of one myocyte and the end of the next, and information is pass across it from cell to cell. These structures are found only in cardiac muscle.
The ID's are vital to normal myocyte function and understanding how they are put together is important. In these special areas there's a considerable degree of
interdigitation of myocyte plasma membrane, but there is no actual
fusion of cytoplasm. The specialized apparatus of the ID allows myocytes to act as
if they were a true syncytium there is in fact no actual cytoplasmic
continuity; these are distinct and individual cells.
The true nature of the ID wasn't fully understood until
the transmission electron microscope was available for their study. This
instrument revealed that ID's are physically held together by large numbers
of desmosomes, with gap junctions between the myocytes forming a region
that permits electrical communication in the form of ion fluxes. This flux
maintains the coordination of the waves of contraction between one cell and the
next.
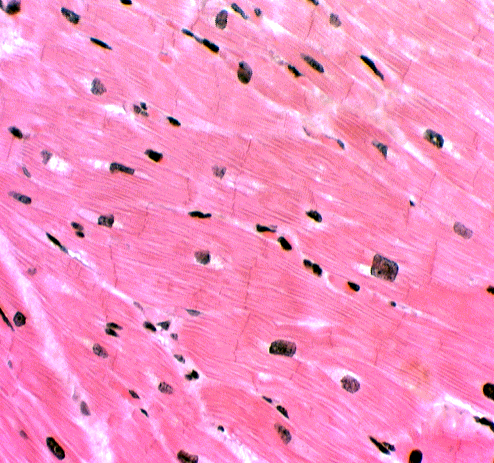 Intercalated discs are readily seen on slide 525, in regions where the myocardium is cut longitudinally (they won't be visible in cross sections). Since the ID's are located at
the ends of the cell, and since that's where the first and last sarcomeres of
any given myofibril are, you can think of the ID as a sort of thickened
"terminal Z-line."
Intercalated discs are readily seen on slide 525, in regions where the myocardium is cut longitudinally (they won't be visible in cross sections). Since the ID's are located at
the ends of the cell, and since that's where the first and last sarcomeres of
any given myofibril are, you can think of the ID as a sort of thickened
"terminal Z-line."
ID's are just barely at the limit of visibility in a light
microscope, and it may help to defocus your substage condenser a bit. They'll
be seen as fuzzy lines running across the width of a row of cells. The presence
of ID's is absolutely diagnostic for cardiac muscle; no other tissue has such
structures.
Laterally there are
no points of fusion or adherence between cells that resemble the ID's. They're only at the ends of the cells. The ID acts as a communication site as well as an adherence structure between two myocytes. Gap junctions between the two myocytes take care of the communication: the ID is a site of lowered electrical resistance, so that passage of ions through the gap junctions into the intercellular space keeps adjacent cells "informed" about what's happening. The coordination of the heartbeat and the transmission of the contraction signal from one cell to the next is dependent on the integrity and proper functioning of the ID's gap junctions. Cell-to-cell adherence is maintained here (as at other sites) by numerous desmosomes. These are scattered along the length of each ID.
Endocardium and Epicardium
 Examine the lining of the chambers on slide 616. The entire cardiovascular
system, including all chambers of the heart and all blood vessels, is lined
with a simple squamous epithelial covering. This lining is among the most
metabolically active tissues in the body. In blood vessels, this is (by
convention) called "endothelium," but in the heart the special term endocardium is used instead. (There isn't a nickel's worth of difference between them, but
the terminology is so entrenched it could never be eradicated.) In any event,
"endocardium" is an apt name because it's literally "inside the
heart." The endocardium covers the valve cusps, too, as you can easily
verify on this slide.
Examine the lining of the chambers on slide 616. The entire cardiovascular
system, including all chambers of the heart and all blood vessels, is lined
with a simple squamous epithelial covering. This lining is among the most
metabolically active tissues in the body. In blood vessels, this is (by
convention) called "endothelium," but in the heart the special term endocardium is used instead. (There isn't a nickel's worth of difference between them, but
the terminology is so entrenched it could never be eradicated.) In any event,
"endocardium" is an apt name because it's literally "inside the
heart." The endocardium covers the valve cusps, too, as you can easily
verify on this slide. 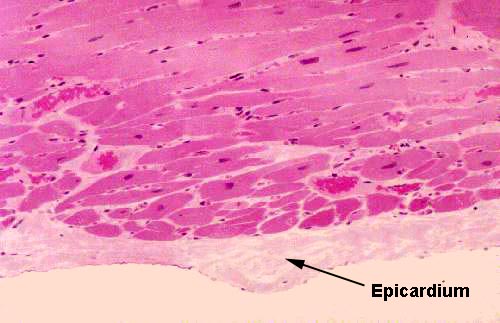 If you traverse the thickness of the myocardium and examine the outer surface of the wall, you will see a similar (though less well defined) layer of
simple squamous epithelium. Again, by convention, this has a special name; it's
the epicardium, which translates as "upon the heart." Anatomically,
this is the visceral layer of the pericardial sac.
If you traverse the thickness of the myocardium and examine the outer surface of the wall, you will see a similar (though less well defined) layer of
simple squamous epithelium. Again, by convention, this has a special name; it's
the epicardium, which translates as "upon the heart." Anatomically,
this is the visceral layer of the pericardial sac.
The epicardium is the inner portion of the CT envelope that surrounds the heart. It corresponds to the peritoneum of the abdominal cavity in origin (from the mesoderm lining the embryonic coelom) and function (to allow the heart to move freely in its cavity without adhesion to surrounding structures). Here you see it as a thin serous membrane overlying the outer surface of the organ. A stain for connective tissue would reveal a very delicate sub-epicardial CT layer, mainly reticular fibers.
Purkinje Fibers
Slide 582 is another chunk of myocardium. Slide 582A is the same thing, but stained differently. Most of the features of myocardium are visible, but this section also
shows Purkinje fibers (Johannes Evangelista von Purkinje, 1787-1869, a Czech
anatomist, physiologist and microscopist). These fibers are part of the
conducting system of the heart.
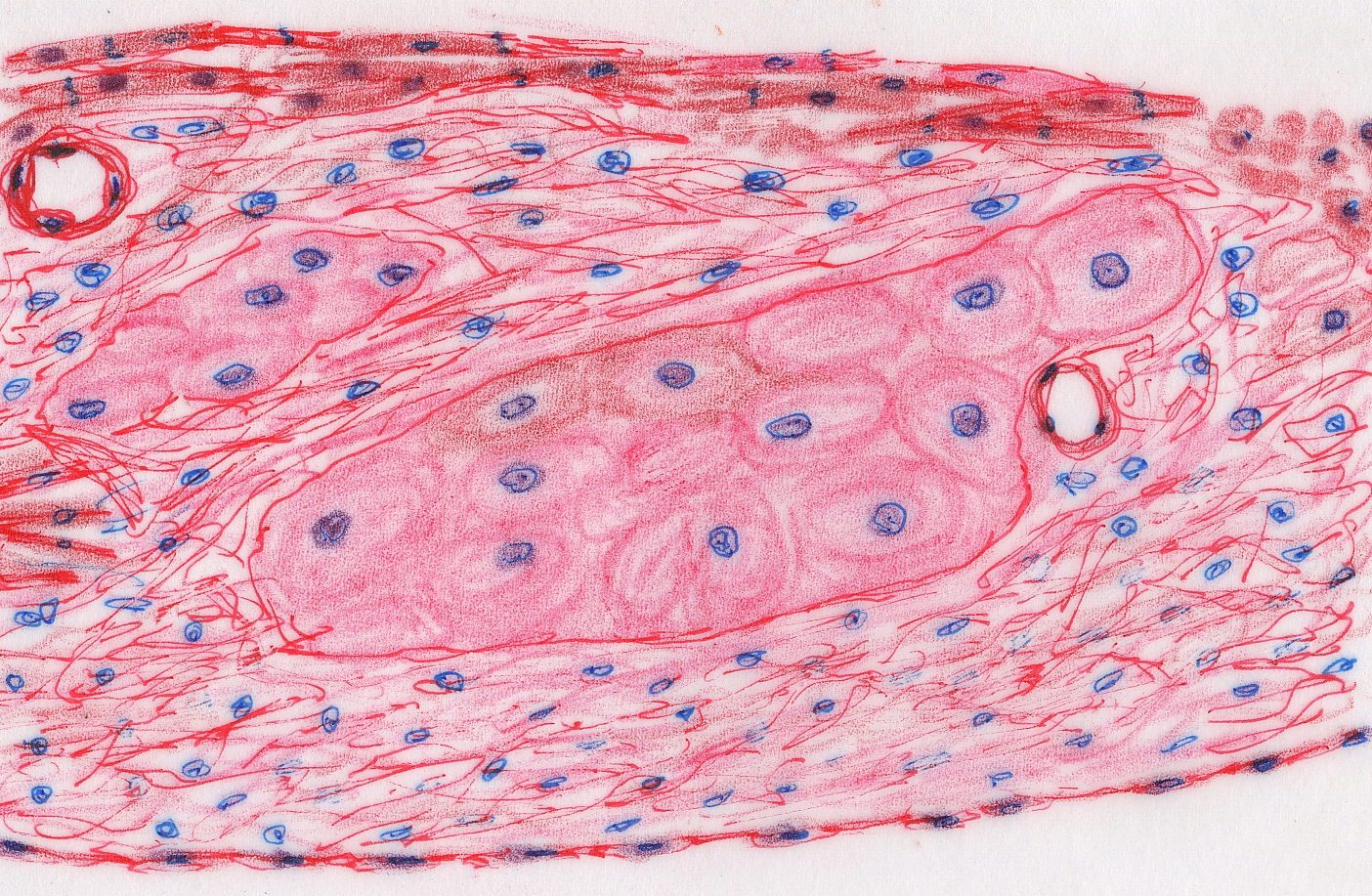
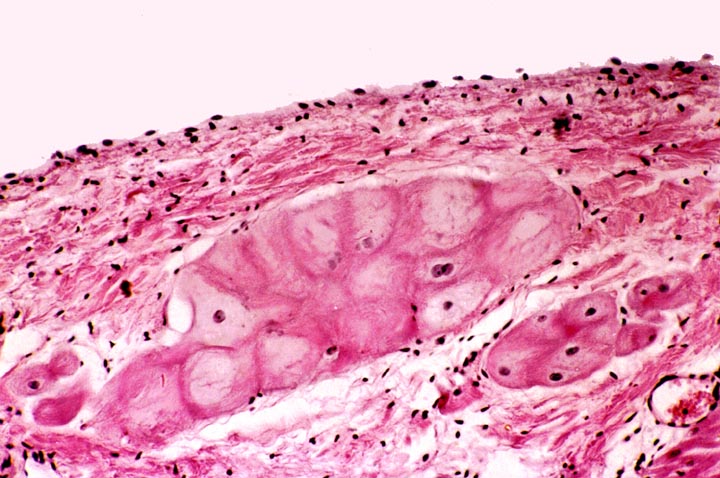
The conducting system transmits the signal to contract from its origin in the "pacemaker" to the rest of the myocardium. The Purkinje fibers are
not neurons. They're specialized muscle cells, and they look like it. They can
be distinguished from the myocardium by their size (they're much larger than
regular myocytes) and by their staining (they tend to be more lightly stained
than true myocardium). With the PAS stain they're strongly positive, since they
contain large amounts of glycogen.

Purkinje fibers are not contractile, at least not to any significant extent. They contribute nothing to the force generation of myocardium. The cells of the Purkinje fibers are organized into several tracts that lead away from the atrioventricular node and form nerve-like structures, but they aren't nerves. There's a bundle along both sides of the septum, one distributing to the right ventricle and one to the left ventricle. The Purkinje fibers are larger and less well stained than the ordinary contractile myocardium because of the relatively protein-poor cytoplasm, which doesn't take up the eosin stain very well. Hence the fibers look pale and washed out in comparison to the protein-packed myocytes. Purkinje fibers do have some of the other features of cardiac myocytes, including a centrally-located single nucleus, and even intercalated disks.
But despite
their strictly conductive function, they are muscle cell derivatives, so you're
quite likely to see striations and other indications of "contractility" if you examine them at higher power, especially in slide 582A. There should be
two areas of Purkinje fibers in your slide, one at each edge of the section. In
this slide also note the ramifications of the muscle, and the intercalated
discs.
The Purkinje fibers ramify into the mass of ventricular myocardium, and from their termini the signal is spread through the ID system. Recall that all muscle tissue is "excitable" and this property can be used to carry information. That's what's happened here. What is a secondary function in contractile myocytes has been transmuted into the main role of these not-quite-muscle cells. The fibers run through the sub-endothelial connective tissue; they can't generate any force because internally they have only a few disorganized actin and myosin fibrils, nothing like the very regular arrays of sarcomeres in the contractile cells.
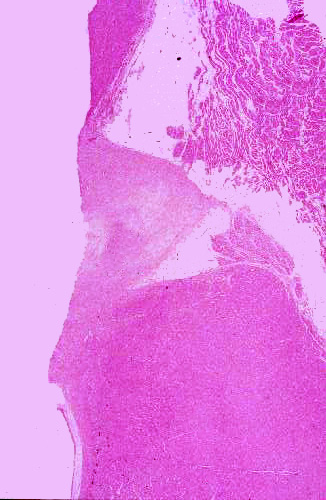 Slide 96, shown at left, is interesting because it contains the atrioventricular node, the beginning of the Purkinje fiber conduction apparatus.
Slide 96, shown at left, is interesting because it contains the atrioventricular node, the beginning of the Purkinje fiber conduction apparatus.
This image is taken with the atrial region at the top of the field. At the upper left is the base of one of the great vessels leading from the heart (probably the aorta); and below that a triangular-shaped region, the trigona fibrosa. The trigona fibrosa is part of the so-called "cardiac skeleton," the dense fibrous CT to which the intercellular collagen network is ultimately anchored. If you look at this structure at high magnification it's very similar in appearance to cartilage. The trigona fibrosa lies between the atrial and ventricular parts of the organ and the fibrous rings of the great vessels are attached to it; so is the top level of the ICN. When force is exerted by contraction of myocardium, this is the "zero point" against which the force is exerted.When contraction occurs, the different directions of muscle tracts impart a compressive, twisting motion to the heart grossly (you will be able to see this in surgery). The motion has been compared to "wringing out" a wet cloth, and it serves the same function: expulsion of the maximum amount of blood per stroke. Contraction of the myocardial cells leads to reduction in volume of the heart chambers, and consequently to ejection of all (or almost all) of the blood therein. Impairment of this action results in decreased efficiency of pumping and eventual failure.
Alongside the trigona, and lying—as its name implies—between the atrium and the ventricle, is the A-V node. It looks like a bit of atrial myocardium, splonked up against the denser and more massive ventricular type. (This slide also gives you a good comparison of the two types of myocardium side by side). This isn't the "pacemaker," (that's the sino-atrial or S-A node) but it receives signals from it and transmits them via the conducting fibers to the mass of ventricular myocardium. Histologically it looks like the rest of the atrial myocardium, and in the absence of the landmarks used here, would be very difficult to identify in a section.
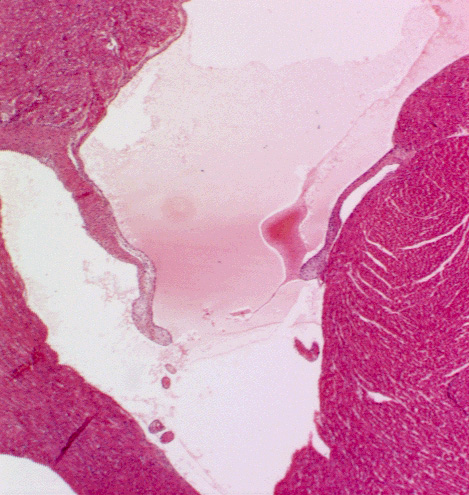
Valves of the Heart
Slide 102 is of a valve cusp. Look at it at low magnification and at high magnification and examine its construction. The cusp of a valve has a core of CT, much of it composed of elastic fibers. There
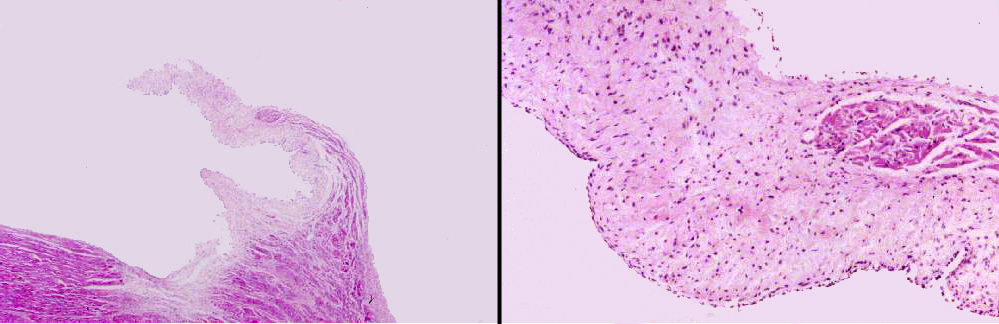
may be some smooth muscle worked into it as well. The outer surfaces, exposed to the blood flow, are covered with the epithelium of the endocardial lining, like the rest of the system. Note that there's an excursion of myocardium into the base of the valve cusp; this terminates before the end of the cusp. It's continuous with the myocardium of the ventricle.
Blood Supply
I hope it hasn't escaped your notice that the entire mass of the myocardium is
shot through with blood vessels of varying sizes and shapes, and that there's
an extensive degree of vascularization. In fact, the heart pumps blood to
itself before it sends any to other parts of the body. The first branches off
the aorta are the coronary arteries of the myocardial circulation. After you
have completed the section on blood vessels you may want to come back and
classify some of these.
Lab Exercise List