Objectives for This Exercise
Objectives for This Exercise
Objectives for This Exercise
Objectives for This Exercise
PRE-RECORDED LECTURES FOR THIS EXERCISE
The digestive system comprises more than just the alimentary canal. In addition to the tubular organs in which actual digestion takes place, there are a number of so-called "accessory glands." That name is really a misnomer; they're vital to normal function and aren't "accessories" to anything. They're every bit as important as the tubular organs are: you'd get along better without a stomach or a fairly good-sized chunk of intestine than without a pancreas. These so-called "accessories" include the liver, the pancreas, and the salivary glands, the first two of which we'll examine in this exercise. Salivary glands will be dealt with in the context of glandular epithelium in Exercise 4.
 The liver is one of the largest organs in mammals, and is the
largest gland by a considerable margin. It's a remarkable machine
with numerous functions, and its structure in many different
taxonomic groups is virtually identical; species differences,
especially within the mammals, are minor.
The liver is one of the largest organs in mammals, and is the
largest gland by a considerable margin. It's a remarkable machine
with numerous functions, and its structure in many different
taxonomic groups is virtually identical; species differences,
especially within the mammals, are minor.
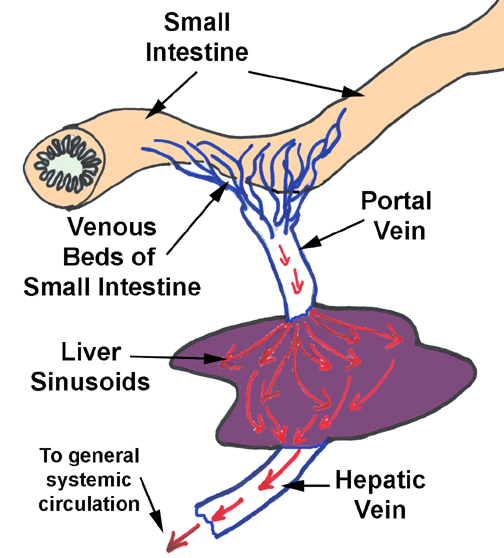
The liver's structure and anatomical placement reflect its function as the site for removal of impurities and for conditioning of the blood. It is, in essence, a large scale "trickle filter" through which the blood must pass before being released to the general circulation. It works in some ways like a sewage treatment plant which purifies wastewater before releasing it back into a river system. The analogy is physically accurate, because the liver slows down the flow of blood by breaking it up into innumerable channels, just as a trickle filter in a wastewater plant passes a large volume of water through sand or gravel to slow the flow for purification to take place. This breakup of the flow is important to normal function.
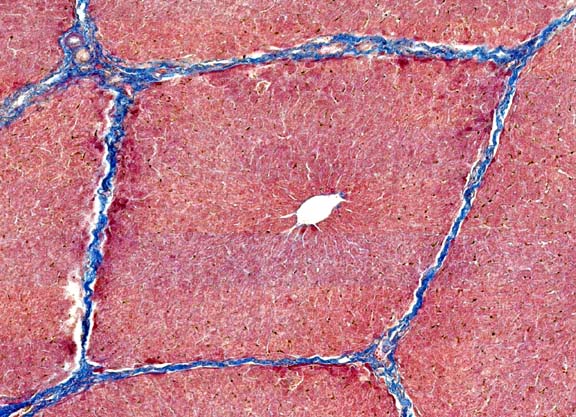
The classic liver lobule is considered by most authorities to be the functional unit of the organ. Each is a roughly hexagonally shaped, three dimensional unit, demarcated by connective tissue and constructed of the parenchymal cells of the liver, hepatocytes, in large numbers.
The image shown here is from the liver of a pig. At moderate magnifications you can pretty easily make out the boundaries of the lobules by observing the direction of radiation of the plates of hepatocytes, but the minuscule amount of CT in most species liver makes it hard to see the lobulation. Pigs are an exception. Like poets, swine are honored more after their death than they are in life, but one of the many virtues of pigs is that their livers have more connective tissue than is true of most other animals; this abundance of CT results in a very 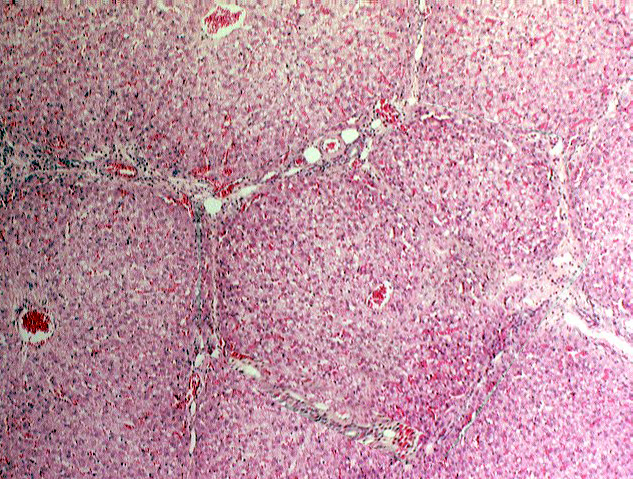 clear demarcation of liver lobules. At this low magnification image the details of the liver's cellular structure aren't easily seen, but the general outline of the lobules can be made out without difficulty. Remember that the lobule is a three dimensional structure, and it projects down into the plane of section, and up out of it, too.
clear demarcation of liver lobules. At this low magnification image the details of the liver's cellular structure aren't easily seen, but the general outline of the lobules can be made out without difficulty. Remember that the lobule is a three dimensional structure, and it projects down into the plane of section, and up out of it, too.
Each lobule is supplied by blood from two sources: a branch of the hepatic artery and a branch of the portal vein. These enter the lobule at its corners, (as part of the portal triad, see below) emptying their blood into the spongy interior of the lobule. What goes in must come out, and blood flowing into the lobule is drained out via the central vein, which marks the beginning of the drainage circulation via the hepatic vein. Central veins from adjacent lobules coalesce to form larger vessels, and eventually all of the drainage leaves the liver via the hepatic vein. Thus blood flow in the lobule is from the periphery to the center.
The lobules (or more correctly, the hepatocytes of which they are made) are the source of the bile, a material important in lipid digestion. Bile is made and transported through a duct system to the periphery of the lobule, where it is drained away via larger ducts. Thus bile flow is from the center to the periphery, exactly the opposite from the blood flow.
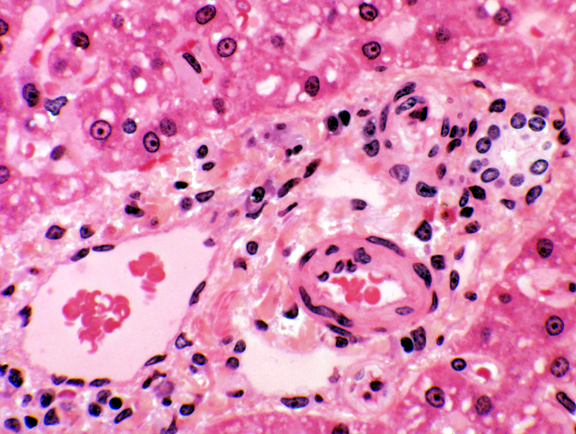 At each corner of the liver lobule, you should be able to make out
a portal canal or hepatic triad. These classically include
at least one each of the following elements: 1) a branch of the
hepatic artery; 2) a branch of the portal vein; and 3)
a bile ductule. There's often a small lymphatic vessel present as well. These vessels are running through the CT stroma which separates the lobule. You won't make out a triad at every corner, but rest
assured that if you could view the lobe in 3-D you would. It's at the triads that blood enters the
lobule and bile leaves.
At each corner of the liver lobule, you should be able to make out
a portal canal or hepatic triad. These classically include
at least one each of the following elements: 1) a branch of the
hepatic artery; 2) a branch of the portal vein; and 3)
a bile ductule. There's often a small lymphatic vessel present as well. These vessels are running through the CT stroma which separates the lobule. You won't make out a triad at every corner, but rest
assured that if you could view the lobe in 3-D you would. It's at the triads that blood enters the
lobule and bile leaves. Blood comes into the lobule from two sources: the portal vein, and the hepatic artery. The venous input is the larger of the two by a substantial margin; that blood is coming to the liver from the capillary beds in the gut (see diagram). The portal vein is carrying blood that's deoxygenated and nutrient-rich. Since the liver is the first place blood reaches, the liver has a chance to detoxify any deleterious materials in the blood before releasing the "cleansed" blood to the general circulation. The arterial supply carries oxygenated blood, and so in the sinusoidal spaces of the liver lobule, oxygenated and deoxygenated blood are mixed together.
The remaining component of the classical triad is the bile ductule. The one labeled here is a nice example, showing how the wall is composed of simple cuboidal epithelium. The bile ductules carry bile produced in the hepatocytes. This small bile ductule one will join with others to form larger and larger vessels, which eventually coalesce into one (or, in some species, two) terminal bile ducts that open into the intestine. The duct system in the liver is set up pretty much like any other exocrine's. It's a compound system, of course. The two ducts shown here are joined together at some point out of the plane of this section, and the smaller one is a tributary of the larger.
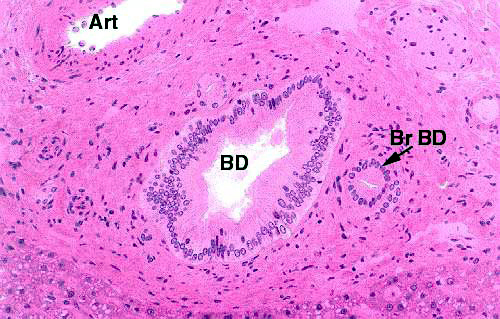 This image shows a large bile duct (BD) and one of its tributaries (Br D). A large artery (Art) is present for comparison. The smaller duct is lined with simple cuboidal to columnar epithelium; the larger duct is big enough that the epithelium is beginning to stratify, and a stratified columnar epithelium can frequently be seen in ducts that size and larger. There's an extensive supporting investment of CT around both of the ducts.
This image shows a large bile duct (BD) and one of its tributaries (Br D). A large artery (Art) is present for comparison. The smaller duct is lined with simple cuboidal to columnar epithelium; the larger duct is big enough that the epithelium is beginning to stratify, and a stratified columnar epithelium can frequently be seen in ducts that size and larger. There's an extensive supporting investment of CT around both of the ducts.
The "triad" might better be called a "tetrad" rather than a triad, because there's usually a lymphatic vessel in the same location. Lymphatic vessels drain off the liquid blood plasma that's released into the intercellular space by flowing blood, and they too are found in the corners of the lobule. They look like very thin walled veins, but—as you can see here—unlike veins they never contain erythrocytes under ordinary circumstances.
 Within the limits that define each lobule, the hepatocytes are arrayed in long rows, and each lobule is organized around a central vein.
Within the limits that define each lobule, the hepatocytes are arrayed in long rows, and each lobule is organized around a central vein. 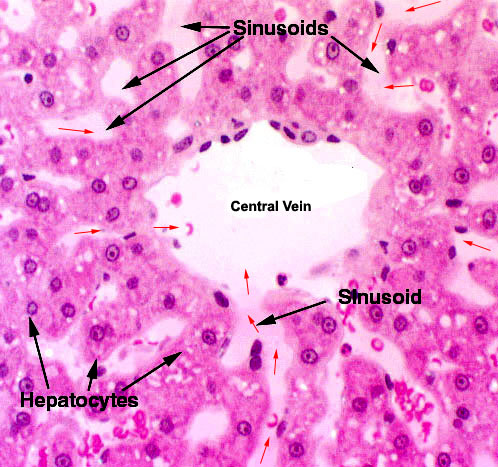 from the arterial and venous input to the liver lobule. You can see erythrocytes in the spaces between the plates of hepatocytes and in the central vein as well.
from the arterial and venous input to the liver lobule. You can see erythrocytes in the spaces between the plates of hepatocytes and in the central vein as well. The sinusoids and the central vein are lined with endothelium (they're blood spaces, after all) and in this image you can see the endothelial cells in the central vein quite easily. This lining is discontinuous. That is, there are places where hepatocytes are "naked" and exposed directly to the flow of blood past them. This is important to liver function as it gives the hepatocytes access to the blood for modification of its makeup. The discontinuity is much greater in the sinusoids—where the real action of the liver goes on—than it is in the veins.
Use slide 123, which has both liver and gall bladder on it, to see the various cell types of the liver; slide #34 may also have some liver present. Both of these are plastic sections stained with H&E, and have very good definition of the structures and cell types described below. Other slides with special stains are noted below, where appropriate.
The archetypical cells of liver, and by far the most numerous cell type, are the hepatocytes of which numerous examples are shown in the images above. Plates of hepatocytes are usually one cell thick. Hepatocytes are fairly uniform in size, and perhaps as many as 25% of them are binucleated.
The hepatocyte is the cell which enables the liver to do all of its tricks of synthesis, storage, and secretion. They make bile, sequester and release carbohydrates, and produce the amazing varieties of protein products for which the liver is responsible.
Glycogen and Lipid Inclusions in Hepatocytes
 The liver has as one of its many functions the responsibility for
maintaining blood glucose levels. To this end, the hepatocytes take
up and store excess glucose as a polymeric derivative,
glycogen. Glycogen can, when needed, be dismantled back into
its constituent glucose moieties by the tireless and faithful
hepatocytes, and put into the blood flowing past them.
The liver has as one of its many functions the responsibility for
maintaining blood glucose levels. To this end, the hepatocytes take
up and store excess glucose as a polymeric derivative,
glycogen. Glycogen can, when needed, be dismantled back into
its constituent glucose moieties by the tireless and faithful
hepatocytes, and put into the blood flowing past them.
Glycogen is PAS positive: the image at right (from slide 500) shows aggregations of it in the hepatocytes, at high (1000x) magnification. It's the coarse magenta material. The boundaries of the hepatocytes and the ordinary architecture of the lobule aren't so obvious in this image as they are in a routine H&E preparation, but this is, in fact, normal liver.
Slides 500 and 501 should be examined as a set. Both stained with the PAS reaction. We've seen slide 500; 501 is a control which before staining was treated with diastase enzyme. Diastase will break down any glycogen present, and hence the staining reaction is abolished. The use of enzymes of known specificities to establish controlled reactions is a basic technique in histochemistry, and this is as nice an example of how it's done as you'll ever find.
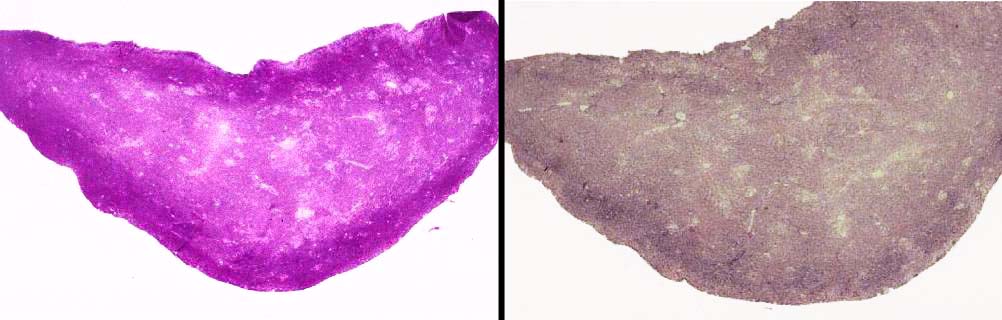
On the left is a section of liver, stained to show glycogen. The deep magenta coloration of the cells is evident. On the right, a section from the same block of tissue was treated with diastase enzyme before the stain was applied. The staining reaction has been abolished. Since diastase is pretty specific for certain carbohydrates, it's possible using controls like this to make some conclusions about the chemical nature of the material being stained. In this case, it's glycogen.
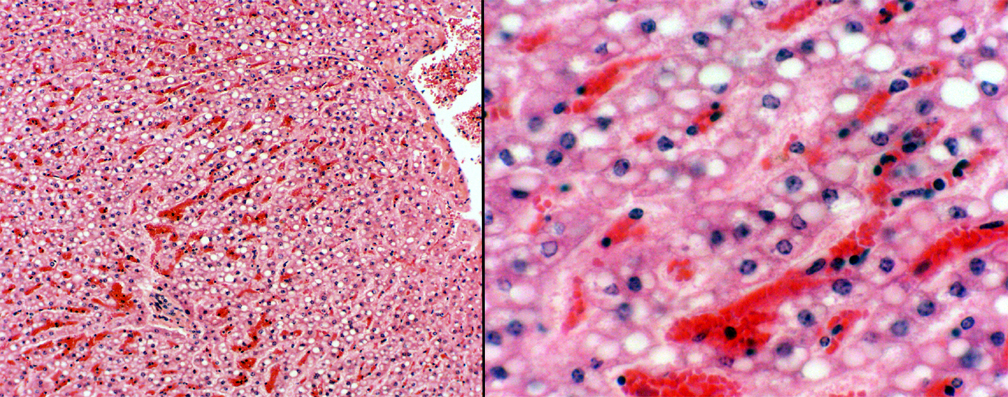
Hepatocytes are a temporary storage depots for lipids,
which they later process into something else. After eating,
especially after eating a lipid rich meal, the liver can be seen to
become gorged with fat droplets in the hepatocytes. This is easily
seen in slide 520. The practice of overindulgence in  spirituous liquors has the same
result, and if you examined the liver of a tipsy reveler on New
Year's Eve, you would see the same sort of thing. What you see above is "reversible fatty change," but "fatty
degeneration" (a less benign and transient condition) can caused by a rather too Bacchanalian lifestyle. It can also be cause by some drugs, such as anti-tumor agents. It's one of
the first indications of liver disease. If the gorging,
imbibing, or other insult continues long enough without respite, the degeneration never
heals properly, and permanent loss of liver tissue, accompanied by
scarring or cirrhosis can occur.
spirituous liquors has the same
result, and if you examined the liver of a tipsy reveler on New
Year's Eve, you would see the same sort of thing. What you see above is "reversible fatty change," but "fatty
degeneration" (a less benign and transient condition) can caused by a rather too Bacchanalian lifestyle. It can also be cause by some drugs, such as anti-tumor agents. It's one of
the first indications of liver disease. If the gorging,
imbibing, or other insult continues long enough without respite, the degeneration never
heals properly, and permanent loss of liver tissue, accompanied by
scarring or cirrhosis can occur.
Occasionally the induction of fatty degeneration is deliberate, at least in animals (we'll regard human drunkards as not having the intention of destroying their livers, whatever the actual outcome is). To produce the delicacy of paté de foie gras, geese are force-fed grain several times daily, by actually holding open their beaks and stuffing it into their gullets with a funnel and stick. The result of this practice—which is illegal in some places—is a heavily fat infiltrated liver.
The sinusoids of the liver—as is true of blood channels in general—are lined by squamous endothelial cells. These are extremely thin and stretched out, laid flat along the tops of the hepatocytes; hence you will be viewing their cut edges. They'll be difficult to see unless you find a place where a nucleus has been sectioned. The lining of the sinusoids is discontinuous, and there are many places where hepatocytes are "naked" and exposed directly to the flow of blood over them. This has obvious implications for an organ that modifies the blood in so many ways as the liver does.
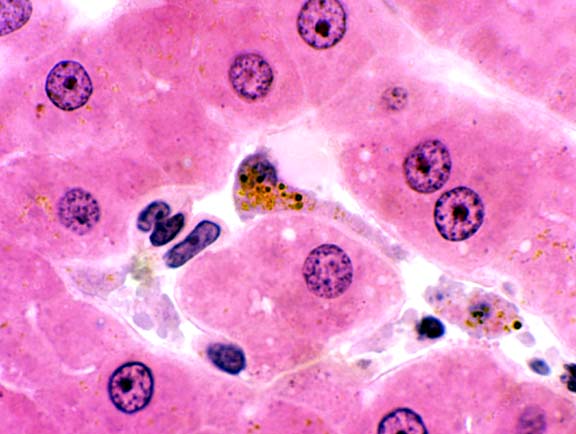 Sitting on top of the hepatocytes and protruding into the sinusoids you'll find another
cell type characteristic of liver. These are the Kupffer cells (Karl Wolfgang von
Kupffer, 1829-1902, a German anatomist). They're phagocytic cells, derived (as are other
examples of macrophages) from the monocyte cell line. These liver-resident macrophages engulf
particulate matter that passes through the sinusoids (especially
bacteria and senescent blood cells).
Sitting on top of the hepatocytes and protruding into the sinusoids you'll find another
cell type characteristic of liver. These are the Kupffer cells (Karl Wolfgang von
Kupffer, 1829-1902, a German anatomist). They're phagocytic cells, derived (as are other
examples of macrophages) from the monocyte cell line. These liver-resident macrophages engulf
particulate matter that passes through the sinusoids (especially
bacteria and senescent blood cells).
You should be able to spot them by the presence of brownish pigment particles inside. This pigment is the product of cell breakdown and can be either lipofuscin or hemosiderin.
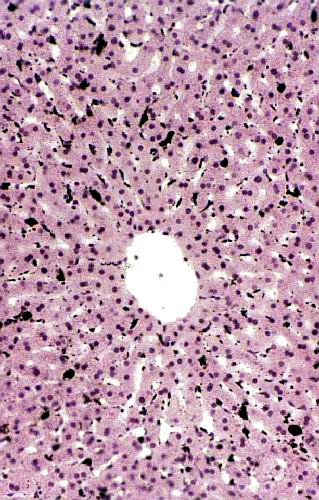 Slide 529 is also worth a look. This low power view shows a liver lobule in a rabbit that's been injected with India ink. The carbon particles of which the ink is composed have been phagocytosed and the Kupffer cells stand out in stark contrast to the hepatocytes around them. The radiating arrangement of the plates of hepatocytes, and the sinusoids between them, are particularly well demonstrated in this slide. (The unusual clarity of the architecture in this preparation is probably due to its having been perfused with fixative; there are no erythrocytes in the sinusoids because they've been "washed out" by pumping fixative in via the hepatic artery and portal vein.) This slide has some historical significance worth a footnote. It's a replication of a classical experiment in the life sciences, one of the pivotal ones that changed the course of modern medicine, by demonstrating that some cells had the capacity for phagocytosis.
Slide 529 is also worth a look. This low power view shows a liver lobule in a rabbit that's been injected with India ink. The carbon particles of which the ink is composed have been phagocytosed and the Kupffer cells stand out in stark contrast to the hepatocytes around them. The radiating arrangement of the plates of hepatocytes, and the sinusoids between them, are particularly well demonstrated in this slide. (The unusual clarity of the architecture in this preparation is probably due to its having been perfused with fixative; there are no erythrocytes in the sinusoids because they've been "washed out" by pumping fixative in via the hepatic artery and portal vein.) This slide has some historical significance worth a footnote. It's a replication of a classical experiment in the life sciences, one of the pivotal ones that changed the course of modern medicine, by demonstrating that some cells had the capacity for phagocytosis.
At higher magnification individual Kupffer cells can be made out pretty well. Note also the relationship of the sinusoids to the central vein, and the fairly large number of binucleated hepatocytes that are present in the plates. The Kupffer cells are literally in position to scarf up things that come by. It's physically located in the sinusoid, separated by a small gap from underlying hepatocytes. This is the Space of Disse (Josef Disse, 1852-1912, a German anatomist).
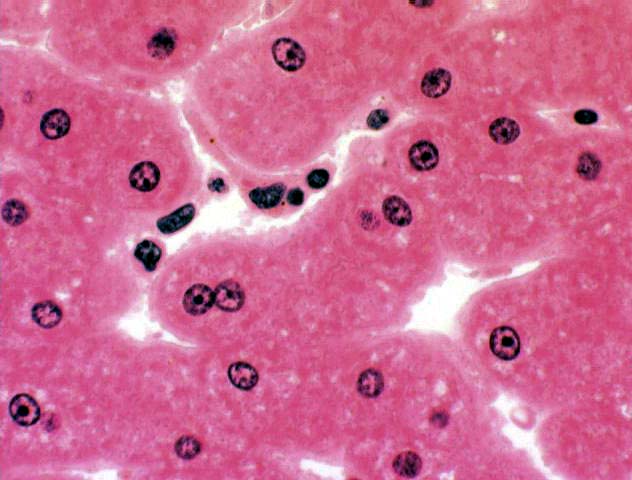 The space of Disse was a subject of considerable controversy before about 1950 as many people regarded it as an artifact of preparation and believed that the Kupffer cells (or, alternatively, the endothelial cells if there wasn't a Kupffer cell around) weren't separated from the hepatocytes. The electron microscope settled that question: the space is real. Transmission electron micrographs even show short microvilli on the surface of the hepatocyte that project into the space of Disse.
The space of Disse was a subject of considerable controversy before about 1950 as many people regarded it as an artifact of preparation and believed that the Kupffer cells (or, alternatively, the endothelial cells if there wasn't a Kupffer cell around) weren't separated from the hepatocytes. The electron microscope settled that question: the space is real. Transmission electron micrographs even show short microvilli on the surface of the hepatocyte that project into the space of Disse.
I have long maintained that the gall bladder is one of the least interesting and most dispensable organs any mammal can have; indeed, many mammals (rats and horses, for example) don't have a gall bladder at all. Its function is to store bile produced in the liver until such time as that bile is needed for the emulsification of fats in the gut lumen. The muscular coat of the gall bladder contracts and injects bile into the intestine. If a gall bladder is surgically removed, about the only serious consequence may be the need to limit the intake of fatty foods. Personally I would regard the loss of French fries and ice cream from my daily life as a horrible tragedy. However, I've known people who've undergone this surgery without any diminution of their ability to eat ice cream, so there's always hope.
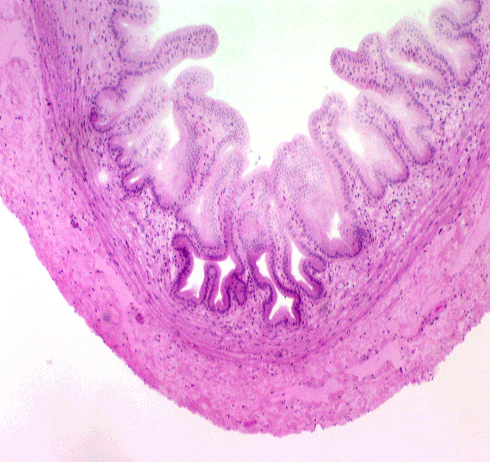
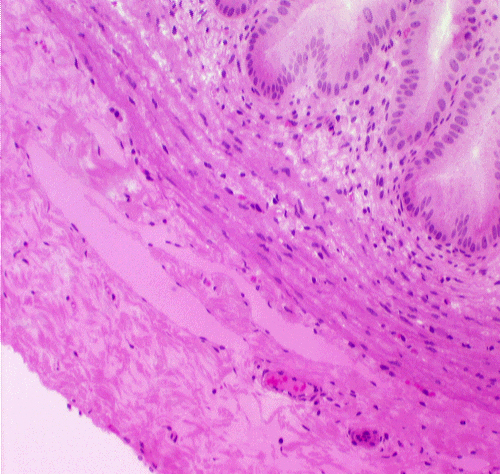 The gall bladder, seen on slide 123, is a tubular organ that fits the typical pattern. Its tunica mucosa is a
very regular simple columnar epithelium, with no goblet cells or
glands. The deep folds in the bottom of the mucosa, which are often
cut in cross section, sometimes look like "glands," but they have no
secretory activity and they are really parts of the lumen. There is a scanty lamina propria, and a thin and stringy tunica
muscularis. There is no muscularis mucosae, and that's one of the "landmarks" for this organ. The serosal covering (i.e., visceral peritoneum) that covers the
portion of the organ that's not nestled up to the liver is probably
absent from your slide, having been removed in handling. The gall bladder is only partially covered by a fold of peritoneum; its opposite side is seated in a pocket in the surface of the liver, attached to it by CT. Bile ducts carry bile to the gall bladder for temporary storage.
The gall bladder, seen on slide 123, is a tubular organ that fits the typical pattern. Its tunica mucosa is a
very regular simple columnar epithelium, with no goblet cells or
glands. The deep folds in the bottom of the mucosa, which are often
cut in cross section, sometimes look like "glands," but they have no
secretory activity and they are really parts of the lumen. There is a scanty lamina propria, and a thin and stringy tunica
muscularis. There is no muscularis mucosae, and that's one of the "landmarks" for this organ. The serosal covering (i.e., visceral peritoneum) that covers the
portion of the organ that's not nestled up to the liver is probably
absent from your slide, having been removed in handling. The gall bladder is only partially covered by a fold of peritoneum; its opposite side is seated in a pocket in the surface of the liver, attached to it by CT. Bile ducts carry bile to the gall bladder for temporary storage.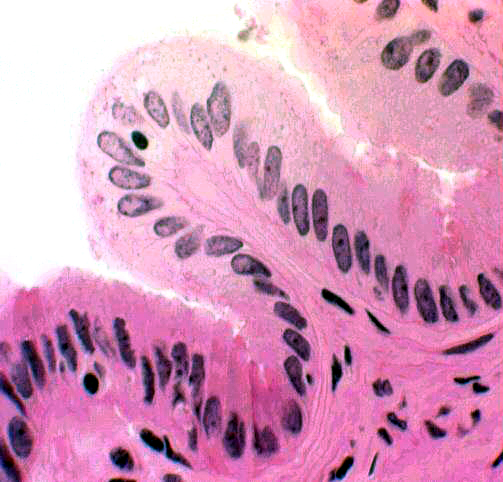 The low magnification image above shows the wall of the gall bladder pretty well. What might are first be taken as villi really aren't. They're folds in the mucosa, permanent rugations like those in the stomach. They vary in height depending on how full the bladder is. When it's distended the folds are lower than you see them here.
The low magnification image above shows the wall of the gall bladder pretty well. What might are first be taken as villi really aren't. They're folds in the mucosa, permanent rugations like those in the stomach. They vary in height depending on how full the bladder is. When it's distended the folds are lower than you see them here.
At higher magnification you can see the nature of the gall bladder epithelium: a rather uninteresting simple columnar form, containing no goblet cells. Though these cells do have a few microvilli on their free surfaces, there is no real brush border. There isn't any muscularis mucosae, and the tunica muscularis is stringy and scanty and shot through with collagen fibers. Since it has only the function of squirting bile into the duct that carries it to the duodenum, it needn't be well-developed.
The pancreas is one organ we can't live without, for a variety of reasons. Mainly because it synthesizes the bulk of the digestive enzymes needed in the intestines; impairment of this function will lead to maldigestion, malabsorption, and eventual death. Inflammation and especially cancer of the pancreas are very serious and often fatal for this reason.
The pancreas is both an exocrine and an endocrine organ; digestive enzymes are conducted via a duct system to the duodenum, and the endocrine products enter the blood directly.
The pancreas is nicely illustrated on slide 34. The bulk of the tissue in the pancreas is exocrine in nature. This is the pancreatic acinar tissue and its associated ducts.
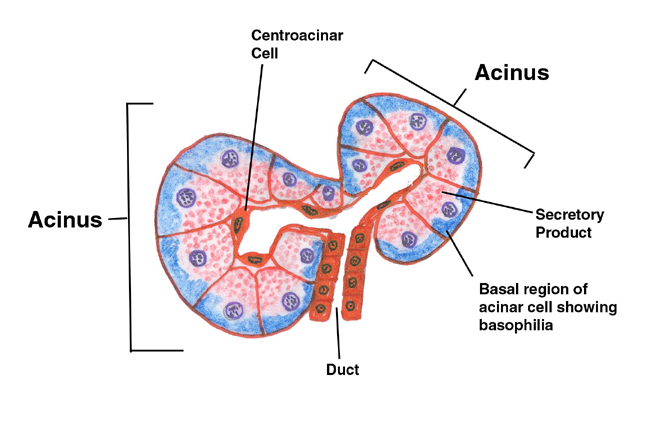 The cells of the acinar tissue are arranged into blind-ended
acini (that's a plural--the singular is "acinus") as in other
glands. The lumen of an acinus is small, but if you have a favorable
section you should be able to make it out. The acinar cells have a
deeply basophilic basal region and an eosinophilic apical area.
The cells of the acinar tissue are arranged into blind-ended
acini (that's a plural--the singular is "acinus") as in other
glands. The lumen of an acinus is small, but if you have a favorable
section you should be able to make it out. The acinar cells have a
deeply basophilic basal region and an eosinophilic apical area.
The concept of the "acinus" is shown here in a diagram. The acinus is the basic secretory unit of the pancreas. It's composed of secretory cells grouped around a lumen into which their secretions are released. The word "acinus" is from the Latin for a "berry" or "grape," and what you're seeing in this sketch is two acini cut along their long axes. In you slide you should be able to resolve the apical region as containing distinct granular material. These zymogen granules are precursor forms of digestive enzymes. The deep basophilia of the basal regions is due to the presence of large amounts of rough endoplasmic reticulum, which binds the hematoxylin of the H&E stain heavily. The secretory product of the acinar cells is stored in vesicles (derived from the Golgi apparatus) and held in the apical cytoplasm until a signal is received to release them. At that time they're brought to the surface of the cell, the granules' membranes fuse with the plasma membrane, and the contents are released into the ductwork by exocytosis. The small duct you see in this sketch will almost immediately join a larger one (see below). The secretory product is released into the ducts in an inactive form and become activated when it reaches the intestine. The reason for this is obvious: the enzymes in it would destroy the pancreas itself before ever reaching the intestine if released in the active form.
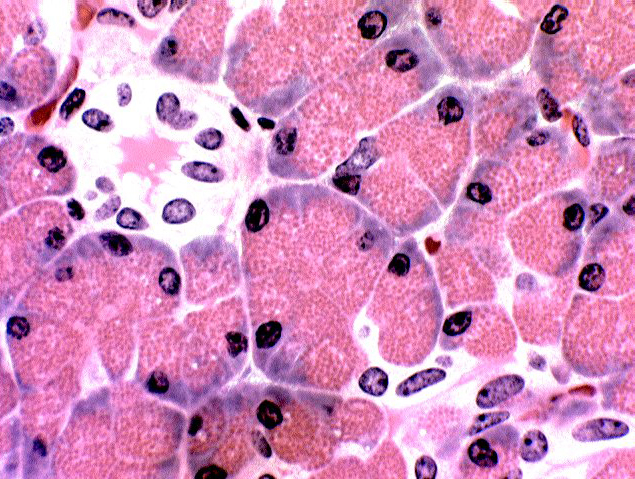 Find a section through the lumen of an acinus, preferably a cross
section. Here you will be able to make out the nucleus of a
centro-acinar cells. This represents the very beginnings of
the duct system. The ducts in the pancreas are not so obvious as
those in salivary glands, but the layout of the glands is
similar.
Find a section through the lumen of an acinus, preferably a cross
section. Here you will be able to make out the nucleus of a
centro-acinar cells. This represents the very beginnings of
the duct system. The ducts in the pancreas are not so obvious as
those in salivary glands, but the layout of the glands is
similar.
The pancreas is a compound acinar gland. The output of the exocrine pancreas is channeled through a series of ducts of increasing caliber. The smallest ones, the intercalated ducts, serve a single acinus; they drain into intralobular ducts, which in turn drain into interlobular ducts, etc., etc. In most animals the entire output of digestive enzymes to the intestine is via a single large pancreatic duct, but an accessory duct is present in some species.
This image shows the most "upstream" end of the system, a secretory acinus and its individual intercalated duct, cut longitudinally. Secretory granules are released by exocytosis and the product begins to flow down the ductwork.
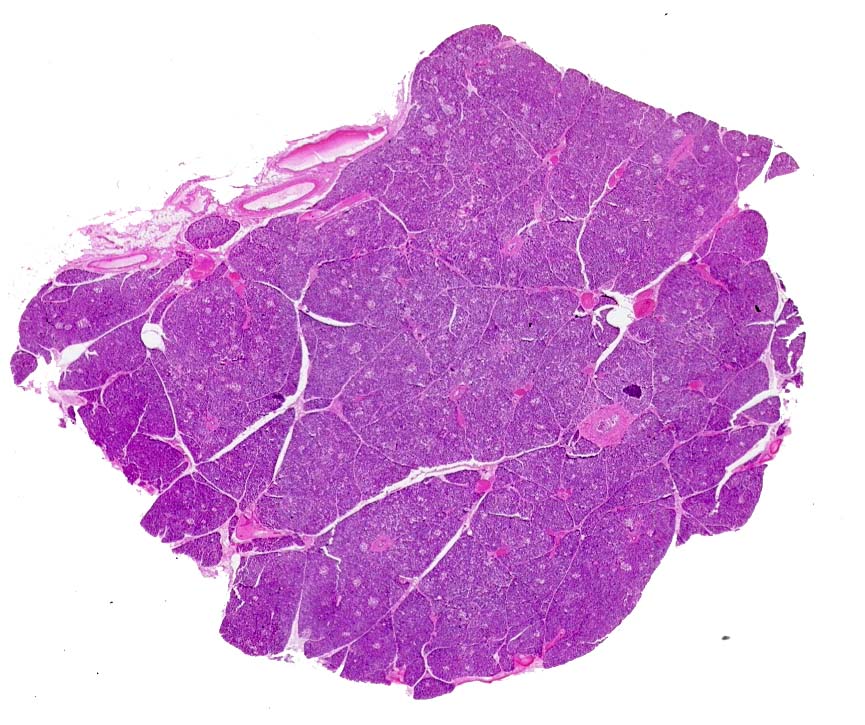
At very low magnification (above) the pancreas can be seen to be extensively lobulated. As with other lobular organs, there's a capsule that surrounds it and CT septae subdivide it into lobes and lobules. 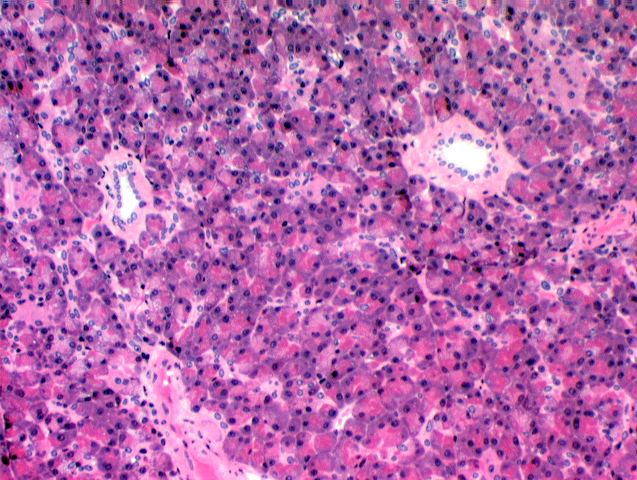 Blood vessels and ducts run through the septae. At medium magnification (left) the larger ducts in the system are easily visible. They're lined with a simple columnar epithelium, but this will stratify as the ducts get large enough. Some of the larger ducts are visible in the image below. As the ducts increase in size their wall becomes more defined and the epithelium of which they're made gets higher; medium sized ducts such as these have cuboidal to columnar epithelium in the wall. Very large ducts will show stratification.
Blood vessels and ducts run through the septae. At medium magnification (left) the larger ducts in the system are easily visible. They're lined with a simple columnar epithelium, but this will stratify as the ducts get large enough. Some of the larger ducts are visible in the image below. As the ducts increase in size their wall becomes more defined and the epithelium of which they're made gets higher; medium sized ducts such as these have cuboidal to columnar epithelium in the wall. Very large ducts will show stratification.
Frequently an acinus is cut in such a way as to reveal only the first few cells in the duct system. These are tucked right up a short way inside the acinus, rather like the stem on a grape. Since they aren't secretory, they'll be stained differently than the secretory cells. The classical histology term for these is the centro-acinar cells. They're just the first cells of the intercalated duct.
 Endocrine Pancreas
Endocrine Pancreas
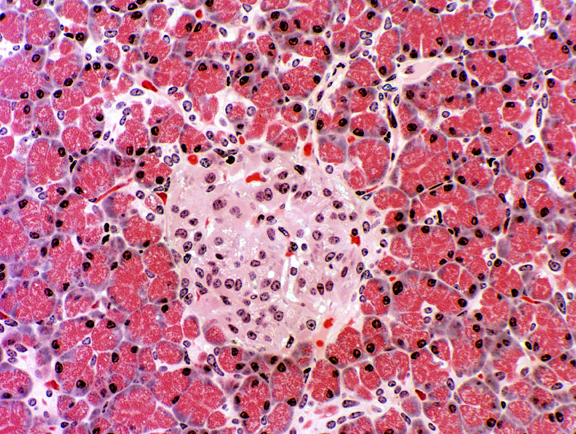 The endocrine portion is the pancreatic islets of
Langerhans (Paul W. Langerhans, 1847-1888, right).
They are regions of lighter staining tissue in the mass of the
exocrine part. The islets are demarcated by a very fine investment of
delicate CT fibrils. They're well vascularized (as you would expect in any endocrine organ) and you can
probably find an erythrocyte or two inside them. The cells produce
the hormones insulin and
glucagon. One of these islets is shown at left. It's literally an "island" of endocrine tissue surrounded by an "ocean" of exocrine acinar pancreas.
The endocrine portion is the pancreatic islets of
Langerhans (Paul W. Langerhans, 1847-1888, right).
They are regions of lighter staining tissue in the mass of the
exocrine part. The islets are demarcated by a very fine investment of
delicate CT fibrils. They're well vascularized (as you would expect in any endocrine organ) and you can
probably find an erythrocyte or two inside them. The cells produce
the hormones insulin and
glucagon. One of these islets is shown at left. It's literally an "island" of endocrine tissue surrounded by an "ocean" of exocrine acinar pancreas.
As is true of all endocrine organs, there isn't any ductwork associated with the islets. They're very well vascularized and the release their products into the blood directly, which carries them to their distant targets. One of the classic experiments done by early investigators was to remove the islet tissue and implant it under the skin. Experimental animals thus treated didn't become diabetic, because when the graft "took" and vascularization occurred, the islet cells could just as easily use the blood vessels of the skin as those elsewhere. in fact, in many non-mammalian species, the endocrine and exocrine functions of the pancreas are physically separate. It's a quirk of evolution that in mammals the two quite different functions are located in the same organ.
Species Variation: Hepatopancreas
It's common in what we featherless bipeds are prone to call "lower" animals to find the pancreas combined with the liver. This is especially true in some fishes. This arrangement is called a hepatopancreas. Examine it and decide which tissue is which. The functions of the cells themselves don't differ from the mammalian system, and at the level of the microscope, the cells of both parts look the same as their mammalian equivalents. (While you're examining this slide, note the nucleated erythrocytes, and compare them to the ones in the exercise on blood!)
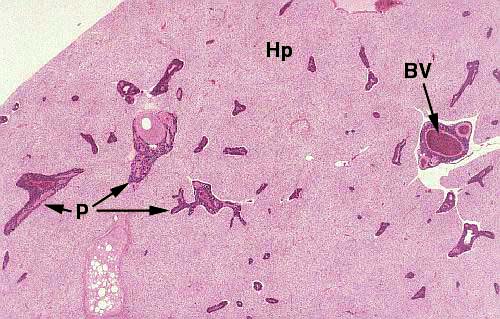 The liver portion (Hp) lacks the distinct lobulation typical of mammals, but the hepatocytes (which comprise most of its bulk) are reasonably similar in appearance and closely similar in function. The really striking thing about this arrangement is the "islands" of pancreatic material scattered throughout the liver. In fact, in fish as in mammals, the liver and the pancreas arise as separate rudiments in embryonic life. The growing pancreas "invades" the liver during development but remains separated from it by CT. Pancreatic tissue (P) proliferates along branches of the hepatic artery (BV) and eventually appears as you see it here, as scattered regions surrounded by liver.
The liver portion (Hp) lacks the distinct lobulation typical of mammals, but the hepatocytes (which comprise most of its bulk) are reasonably similar in appearance and closely similar in function. The really striking thing about this arrangement is the "islands" of pancreatic material scattered throughout the liver. In fact, in fish as in mammals, the liver and the pancreas arise as separate rudiments in embryonic life. The growing pancreas "invades" the liver during development but remains separated from it by CT. Pancreatic tissue (P) proliferates along branches of the hepatic artery (BV) and eventually appears as you see it here, as scattered regions surrounded by liver.
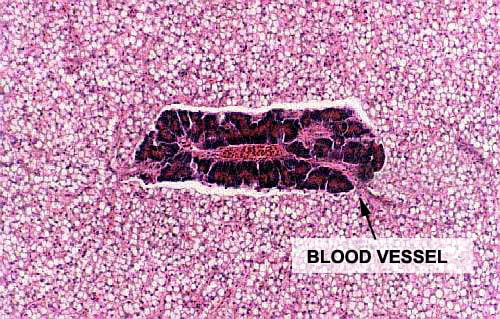 In this image the relationships of the pancreatic tissue to the liver and to the blood vessels are clearly seen. A blood vessel runs right through the "island." The pancreatic cells themselves are virtually indistinguishable from their mammalian counterparts: densely basophilic basal regions and bright eosinophilic secretory granules. There is a duct system into which they empty, though you can't see it in this image.
In this image the relationships of the pancreatic tissue to the liver and to the blood vessels are clearly seen. A blood vessel runs right through the "island." The pancreatic cells themselves are virtually indistinguishable from their mammalian counterparts: densely basophilic basal regions and bright eosinophilic secretory granules. There is a duct system into which they empty, though you can't see it in this image.
Not all fish have a hepatopancreas exactly like this, though most do. Fishes are a very old group in evolutionary terms, by far the oldest group of vertebrates. They have had so many millions of years in a fairly stable environment that adaptive radiation has produced some oddities in their anatomy and physiology that might well have been weeded out in a terrestrial setting. Some fish have their pancreatic tissue formed into discrete organs; some have it scattered as clumps of parenchyma in the mesenteries that support the intestine. There are even some that have carried things to the logical extreme and have their pancreatic tissue scattered diffusely in the mucosal lining of the intestine! But the hepatopancreatic layout seems to have been the dominant theme.