Formed Elements of Blood
The tree of Liberty must be refreshed from time to time with the blood of
patriots and tyrants.
—Thomas Jefferson (1743-1826)
Letter to William S. Smith (1787)
Objectives For This Exercise
CASE VIGNETTE
SLIDES USED IN THIS EXERCISE: VM11, VM12, VM13, H9020, 1004, VM05, 98, VM01
THERE ARE PRE-RECORDED LECTURES FOR THIS EXERCISE
Please go to the listing page for these to download and view them
Formed Elements of Blood
Bone Marrow and Hemopoeisis
Blood Is A Connective Tissue
Blood is classically considered to be one of the connective tissues, since it
has all of the three components which characterize these: fibers, an amorphous
matrix, and cells. The fibrous component is latent in normal blood, and is normally expressed only in
response to injury. The amorphous matrix is blood plasma, a low
viscosity aqueous solution of proteins and some other constituents. In this
laboratory exercise we will consider only the cellular and cell-derived components, i.e., the "formed elements"
of normal circulating blood. The true cells (as distinct from non-cells) found in circulating blood are in
many cases the same cells found in loose CT's, and this is yet another reason
for putting blood in this category of basic tissues.
Wright's Stain
In most of this exercise you'll be using smear preparations made with Wright's stain (named after its
developer, James H. Wright, 1871-1928, an American pathologist). A blood smear is made by putting a drop on a slide and spreading it into a thin film, which is fixed and stained simultaneously. Wright's is an
alcohol based mixture of rosin and methylene blue whose staining reactions are similar (but not identical to) the common H&E stain used on tissue sections. It's sold commercially under various names, the most familiar being "Diff-Quick." The slide is dipped into the first stain jar, rinsed, and then dipped into the second. Then it's air dried and cover-slipped.
Most people develop a mental image of what formed elements of blood "ought" to look like based solely on smears made this way; and specifically, based on smears stained with Wright's solution. Unfortunately, as useful as these smears are, you could hardly find a less congenial environment for the somewhat delicate formed elements. They're held down to the glass of the slide by surface tension, stretched and flattened out like a road-killed possum on Interstate 81. Under these conditions, they tend to look larger than they really are. There are also subtle differences in the colors produced by Wright's stain and the common H&E stain used in sections.
Because it's important to be able to identify cells in sections, I've tried to include examples of the various cell types from both situations. You need to become skilled at spotting blood cell types in sections, and to remember that however useful smears are (and they're very useful) the differences in appearance exist.
Commonalities of Formed Elements and Cells of CT
Most of what we think of as "blood cells" are really cells of connective tissues
in general. Most of them use the bloodstream simply as a transport system,
residing in it only a few hours, or at most a couple of days. Almost any of the
formed elements can normally be found outside the circulatory system,
interspersed among the fibers of irregular CT. Some of the circulating cells
are immature forms, whose true nature (and final morphology) are demonstrated
only after they take up residence in the CT.
Erythrocytes & Platelets
Begin with slide 1004, a smear of normal canine blood. (If you're a cat person, try slide 1001, normal feline blood, instead.) I've put these two formed elements first because they have some things in common: in mammals they're not true cells, but are derived from true cells. They lack a nucleus and organelles and have limited, specific functions.
The most common formed element is the erythrocyte, or "red blood cell" (RBC).
Despite the common term, this is not a cell at all, although it's derived from
true cells. The RBC is the mature stage of development of a cell line in which
the nucleus (present in earlier forms) has been lost.
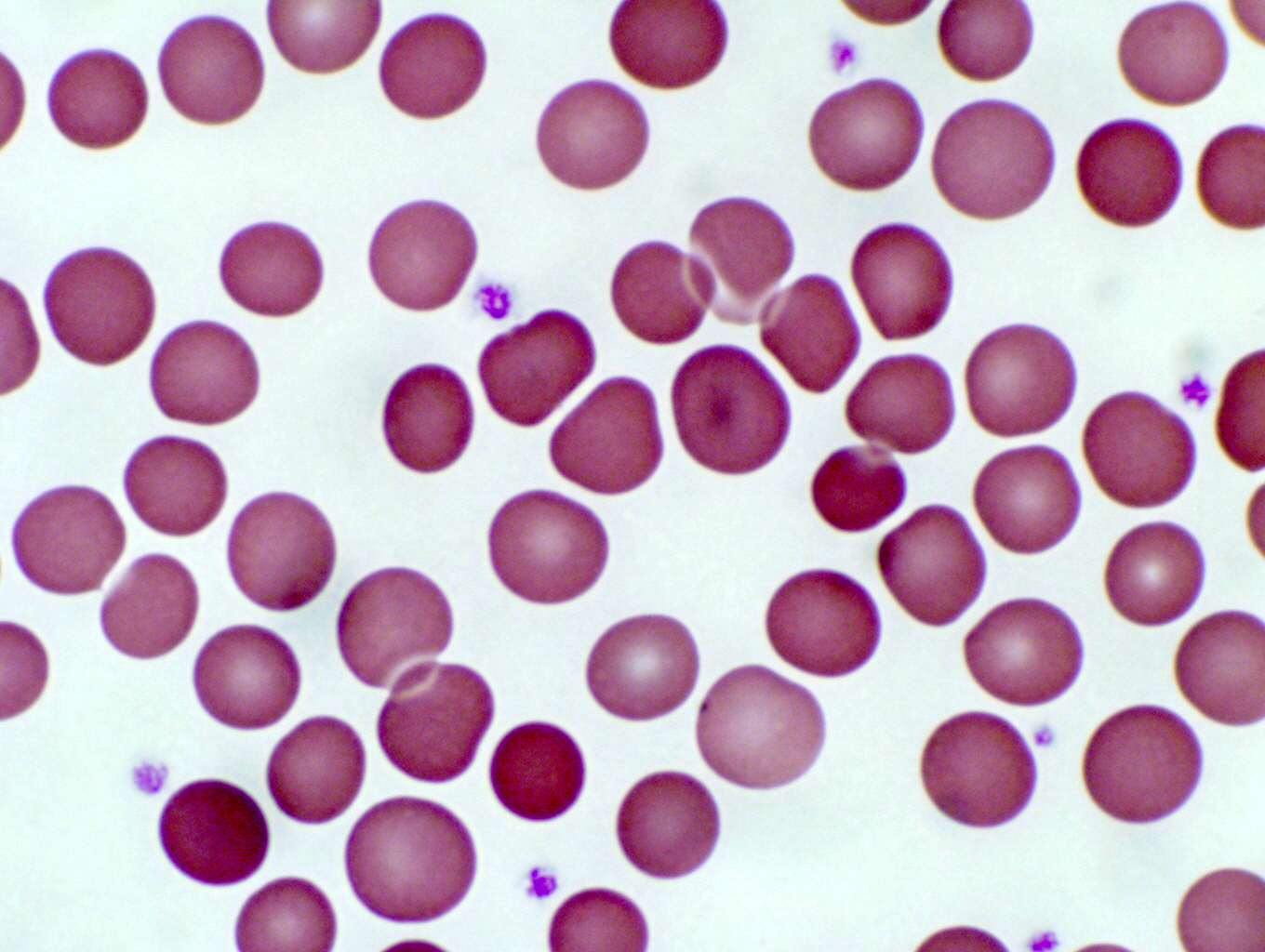
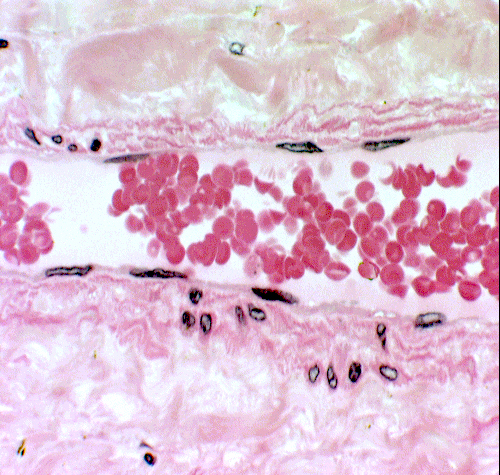 Structurally RBCs are pretty uninteresting: they're merely bags of hemoglobin,
the oxygen transport material. They usually take on the shape of a bi-concave
disc, and have no internal organelles. Even in the EM they are internally
uniform and amorphous. In the image at left there are a few of the millions scattered this slide of a smear. At right, a few hundred are seen in the lumen of a small artery, at slightly lower magnification than in the smear.
Structurally RBCs are pretty uninteresting: they're merely bags of hemoglobin,
the oxygen transport material. They usually take on the shape of a bi-concave
disc, and have no internal organelles. Even in the EM they are internally
uniform and amorphous. In the image at left there are a few of the millions scattered this slide of a smear. At right, a few hundred are seen in the lumen of a small artery, at slightly lower magnification than in the smear.
The morphology of RBCs is pretty much the same in all the common mammalian
species; most of the variations in shape can be accounted for as processing
artifacts and/or the results of disease. One notable exception is in the
camelids (camels, dromedaries, and llamas). These animals have oval RBCs,
rather than the round ones found in all other mammals.
At one time, though, the precursors of erythrocytes were true nucleated cells. These precursors exist in the bone marrow. As the last step of the developmental process, the nucleus is extruded. At that point the fully-formed erythrocyte is normally released into the blood circulation. Under normal circumstances there shouldn't be any of the nucleated precursors in circulation, but sometimes, if there's a high demand for new erythrocytes (as, for example, when there's continuing blood loss) some not-quite-mature cells will be released with nuclei in them. These will later be expelled while in the circulation.
The life span of erythrocytes is limited: about 90-120 days. After that time they develop surface "senescence" markers and get engulfed in the spleen and liver by resident macrophages. The hemoglobin and plasma membranes are broken down and scavenged for re-usable components, and the residue becomes the iron containing pigment hemosiderin.
There's one thing that RBCs are handy for (besides carrying oxygen, of course).
In most mammals they're about 5.5 to 7.5 microns in diameter, and very
uniform in size (as the pictures make clear). They make a handy built-in "size gauge" in tissue sections, since
there are always a few around somewhere in a tissue section. It's worth your time to look for "free" erythrocytes such as the ones above, and use them to get a better idea of the size of things in the other parts of the field.
Platelets
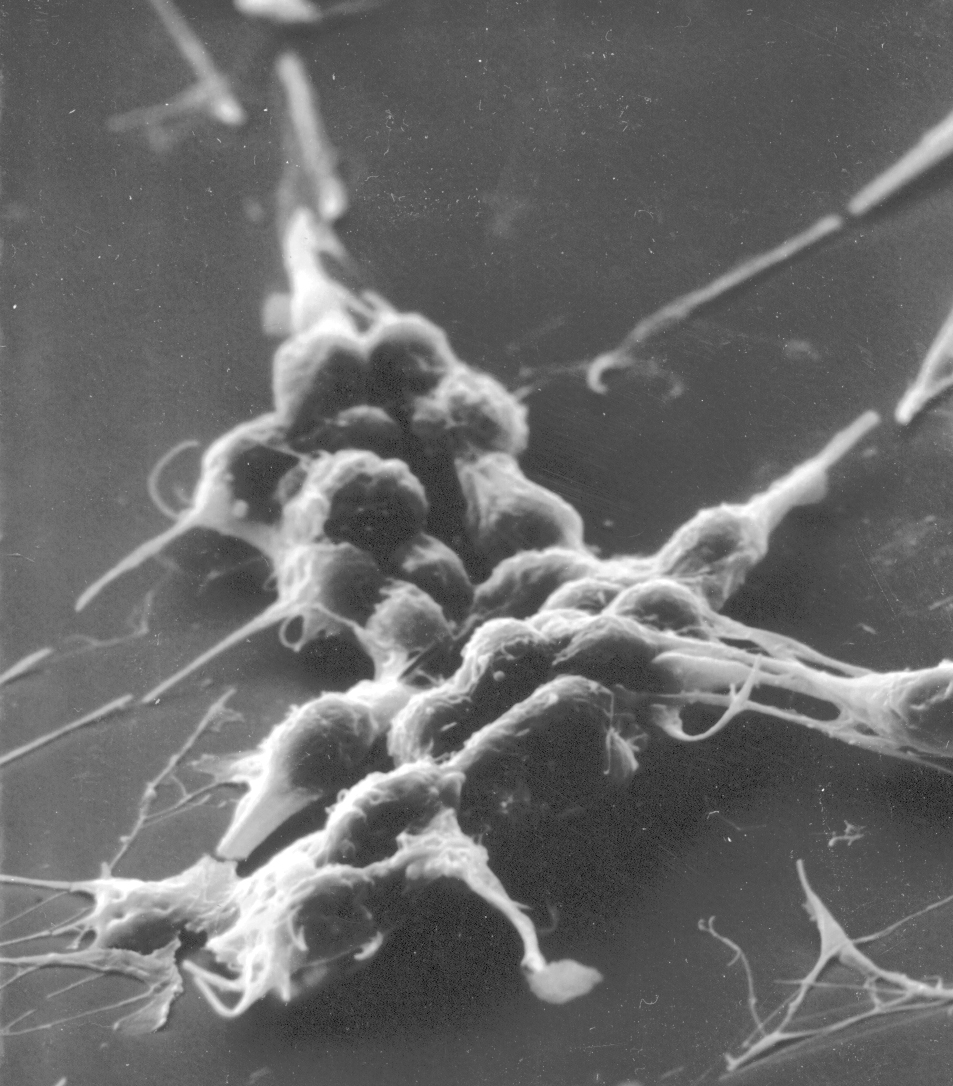 The platelet, like the erythrocyte, isn't a cell at all. Platelets initiate the clotting process. Like the erythrocyte, the platelet was once part of a cell, but in its maturation it loses its true cellularity. In fact, it's nothing but a fragment of a pre-existing precursor cell of the bone marrow a very large megakaryocyte. In the course of its differentiation, the megakaryocyte develops fissures in its cytoplasm, and literally falls apart, the membrane bound cytoplasmic fragments being thenceforth known as platelets. Platelets are little bitty things, only 2 to 4 microns in diameter (about half the size of an RBC), and round to oval in shape. They have a central zone that is slightly basophilic, and a pale, homogeneous periphery. Look for them in the spaces between erythrocytes. The image above shows platelets in a smear: they appear as small fragments with a blue cast to them. They're much smaller than the erythrocytes: maybe 1-3 microns. The scanning electron micrograph at the right is of a group of platelets which have begun to initiate a clot. The stringy material covering them is fibrin, the matured, insoluble fibrillar component of blood.
The platelet, like the erythrocyte, isn't a cell at all. Platelets initiate the clotting process. Like the erythrocyte, the platelet was once part of a cell, but in its maturation it loses its true cellularity. In fact, it's nothing but a fragment of a pre-existing precursor cell of the bone marrow a very large megakaryocyte. In the course of its differentiation, the megakaryocyte develops fissures in its cytoplasm, and literally falls apart, the membrane bound cytoplasmic fragments being thenceforth known as platelets. Platelets are little bitty things, only 2 to 4 microns in diameter (about half the size of an RBC), and round to oval in shape. They have a central zone that is slightly basophilic, and a pale, homogeneous periphery. Look for them in the spaces between erythrocytes. The image above shows platelets in a smear: they appear as small fragments with a blue cast to them. They're much smaller than the erythrocytes: maybe 1-3 microns. The scanning electron micrograph at the right is of a group of platelets which have begun to initiate a clot. The stringy material covering them is fibrin, the matured, insoluble fibrillar component of blood.
Leukocytes
The "white blood cells," or leukocytes, are true cells with nuclei and
organelles. Despite the term "white blood cell," these really aren't "blood" cells at all. They generally function (and in some cases mature) only after leaving the blood compartment for the connective tissue space, and hence they're connective tissue cells. The blood is their means of transit. These cells are capable of recognizing specific sites in the walls of blood vessels, and squeezing through these locations (a process called diapedesis) using a amoeboid form of motion.
They're subdivided into two major categories. Granulocytes have
inclusions in their cytoplasm; agranulocytes don't. The granulocytes
also have lobulated or segmented nuclei; the agranulocytes typically do not.
The principal granulocytes are neutrophils, eosinophils, and basophils.
The agranulocytes are the lymphocytes and the monocytes.
Granulocytes: Neutrophils
 Neutrophils are the most common form of leukocyte in primates, dogs, cats,
and horses. They have other names, frequently encountered in clinical
literature: "polys" or "PMNs" are most common, both abbreviations for
"polymorphonuclear leukocyte." These cells can be identified on a smear by
their size (about 9 to 12 microns diameter) and their characteristically segmented
or lobulated nucleus. The nucleus in its mature form may have 3 to 5
lobes, connected together with fine threads of nuclear material. The granules
in the cytoplasm are usually lightly stained, and may appear to be pink to
purple. This variability of staining has resulted in the name "heterophil"
being applied to these cells. Neutrophils might be termed the "shock troops" of the body's defense against disease. Like World War One infantry, their role is to "do or die" in the Great War Against Infection. When the physical integrity of the body is breached and foreign objects (especially bacteria) manage to enter, neutrophils aggregate at the site in enormous numbers by chemotactic factors released from the damaged tissue. Once there, they fix bayonets, go Over The Top, and charge the invaders, engulfing them and destroying them by lysis. In the course of this heroic action, the neutrophils make The Ultimate Sacrifice and die. Their lysed remnants constitute pus, and a wound that is oozing pus is said to be suppurative, or to "have a suppurative exudate."
Neutrophils are the most common form of leukocyte in primates, dogs, cats,
and horses. They have other names, frequently encountered in clinical
literature: "polys" or "PMNs" are most common, both abbreviations for
"polymorphonuclear leukocyte." These cells can be identified on a smear by
their size (about 9 to 12 microns diameter) and their characteristically segmented
or lobulated nucleus. The nucleus in its mature form may have 3 to 5
lobes, connected together with fine threads of nuclear material. The granules
in the cytoplasm are usually lightly stained, and may appear to be pink to
purple. This variability of staining has resulted in the name "heterophil"
being applied to these cells. Neutrophils might be termed the "shock troops" of the body's defense against disease. Like World War One infantry, their role is to "do or die" in the Great War Against Infection. When the physical integrity of the body is breached and foreign objects (especially bacteria) manage to enter, neutrophils aggregate at the site in enormous numbers by chemotactic factors released from the damaged tissue. Once there, they fix bayonets, go Over The Top, and charge the invaders, engulfing them and destroying them by lysis. In the course of this heroic action, the neutrophils make The Ultimate Sacrifice and die. Their lysed remnants constitute pus, and a wound that is oozing pus is said to be suppurative, or to "have a suppurative exudate."
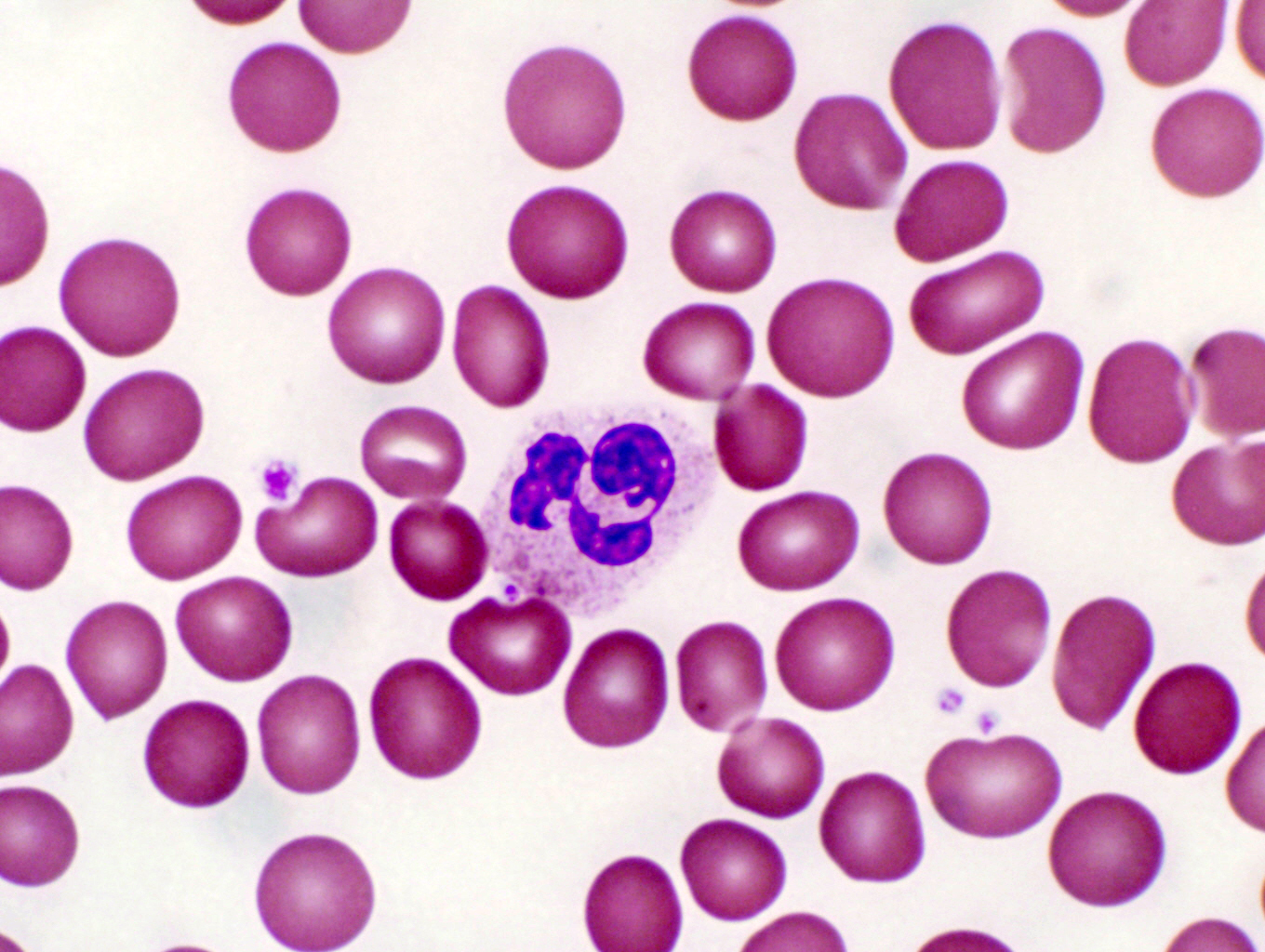 The granules in the neutrophil are lysosomes, and
the neutrophil is a phagocyte, particularly for bacteria. They are commonly
found in the tissues at the site of bacterial invasions. The neutrophils at
these sites engulf and destroy invading bacteria, and then die; their dead and
lysed bodies constitute the bulk of pus at sites of suppurating wounds.
The granules in the neutrophil are lysosomes, and
the neutrophil is a phagocyte, particularly for bacteria. They are commonly
found in the tissues at the site of bacterial invasions. The neutrophils at
these sites engulf and destroy invading bacteria, and then die; their dead and
lysed bodies constitute the bulk of pus at sites of suppurating wounds.
Neutrophils are recognized in smears by their characteristic lack of staining. Although they contain numerous granules (the lysosomes with which they destroy bacteria) these don't pick up much stain and the cytoplasm remains comparatively unstained. Another characteristic is the nuclear morphology. The nuclei of these cells are highly variable in shape and usually (as is this one) extensively lobulated. When a neutrophil has recently released into the blood from the marrow, its nucleus forms one or two lobes, but as the cell matures it develops more and more. All the lobes are connected to each other by thin, threadlike extensions of the nuclear substance, however. There is only one nucleus per cell, though if the connections are very tenuous there may appear to be more.
The one in the smear shown here is from a monkey. Specifically, a female monkey. The extensive lobulation of the nucleus has produced a drumstick shape lobe, a "Barr Body" (named for Murray L. Barr, 1908-1995, a Canadian anatomist) which represents the extra X chromosome in females.
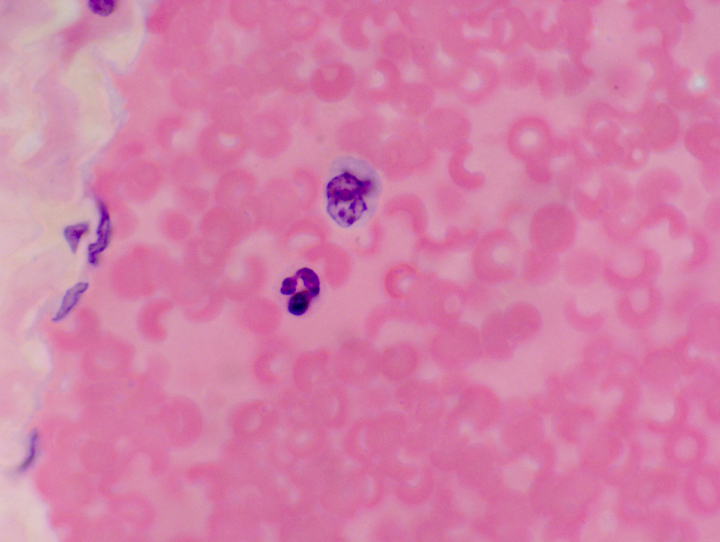 In the circulation, as in the image at left, and when stained with H&E, as here, the neutrophil looks a little different than in smears. Its cytoplasm tends to be slightly eosinophilic, and hence is usually "lost" in the background staining of the protein-rich plasma. The most reliable identification is the multi-lobed nucleus. Typically the granules of neutrophils are very small, and since they're also pretty unreactive to the H&E stain, they usually aren't obvious. The one at left is in a vein, and there's a monocyte next to it, a much larger cell than the neutrophil (see below). And again, in circulation the cell appears to be smaller than it does in smears because it isn't flattened out.
In the circulation, as in the image at left, and when stained with H&E, as here, the neutrophil looks a little different than in smears. Its cytoplasm tends to be slightly eosinophilic, and hence is usually "lost" in the background staining of the protein-rich plasma. The most reliable identification is the multi-lobed nucleus. Typically the granules of neutrophils are very small, and since they're also pretty unreactive to the H&E stain, they usually aren't obvious. The one at left is in a vein, and there's a monocyte next to it, a much larger cell than the neutrophil (see below). And again, in circulation the cell appears to be smaller than it does in smears because it isn't flattened out.
The size of neutrophils in sections ) especially in the very thin plastic section shown here) may seem to vary quite a bit, but that's an illusion of preparation. Remember, these are spherical objects. In a section like this one, which is perhaps 2 microns thick, it's possible to pass the section plane through it at any "latitude" of its 12 micron diameter, so it may appear smaller than it actually is at its maximum width.
Another point to keep in mind is that the lobulated nucleus of this cell can be oriented in any way at all with respect to that section plane. In this image it's easy to see that the lobules are connected, but if the cell had happened to be turned so that the section plane passed through it differently, you might well have seen "unconnected" lobules, easily misinterpreted as multi-nucleation.
Granulocytes: Eosinophils
Eosinophils are much less common than neutrophils. They're the same size
or even slightly larger (10 to 14 microns) and their cytoplasmic granules are
strongly eosinophilic, staining a bright orange-pink color. Typically the
nucleus is bi-lobed, not multi-lobed as in the neutrophils. There's some species variation in eosinophils. The granules are exceptionally
large in horses (slide 1006) and it would be worth your while to compare the
"typical" eosinophils from dogs and cats with those of the horse. Their
function isn't completely understood, but it's known that eosinophils are
involved in allergic responses.

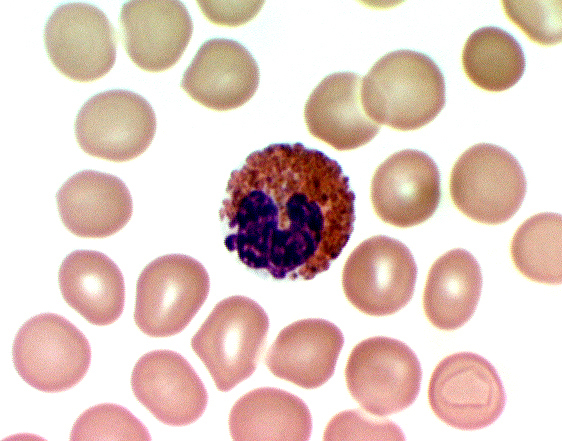 Like the granules of neutrophils, the eosinophil's granules contain lytic enzymes, but they're much larger than those of neutrophils, and they stain pinkish red in Wright's smears. The staining reaction is the most characteristic means of identifying these cells.
Like the granules of neutrophils, the eosinophil's granules contain lytic enzymes, but they're much larger than those of neutrophils, and they stain pinkish red in Wright's smears. The staining reaction is the most characteristic means of identifying these cells.
Another useful clue is the nuclear morphology. Eosinophils never show the extensive nuclear lobulation typical of the neutrophil, but typically have a bi-lobed nucleus. Because the granules can be so prominent, the lobation of the nucleus can sometimes be difficult to see.
 Eosinophils have been proposed to engage in phagocytosis, but not of particulate antigens: rather to engulf and destroy the precipitated antigen-antibody complexes produced in hum orally based immune reactions. It's known that the eosinophil counters the action of many mast cell mediators by
secreting degradative enzymes, including histaminase and aryl sulphatase,
which destroy histamine and leukotrienes respectively. In helminthic
infestation, major basic protein and eosinophil cationic protein released from
the eosinophil are probably beneficial in promoting elimination of the parasites. An elevated eosinophil count is typical in allergic reactions and in parasitic infestations, which support both these putative functions. Eosinophils chemotactically aggregate in large numbers to sites where antigen-antibody complexes are found and where the host is attempting to deal with foreign bodies. They're found easily in the connective tissue below a wheal, the raised "bump" that results at the site of an insect bite. The strong immune response such a bite elicits brings them in, and a section taken through a wheal will have thousands of eosinophils evident. Here's an example of such an eosinophilic exudate. Note the very bright and obvious staining of these cells, as well as the nuclear configuration. In an H&E stain, these could easily be mistaken for mast cells at this low magnification, but such huge aggregations are never seen in normal situations. Finding eosinophils in circulating blood is, frankly, a matter of dumb luck. They aren't anywhere near as numerous as neutrophils (they make up about 0.5-3.0 % of all white blood cells, depending on the species) so spotting one is usually an accident.
Eosinophils have been proposed to engage in phagocytosis, but not of particulate antigens: rather to engulf and destroy the precipitated antigen-antibody complexes produced in hum orally based immune reactions. It's known that the eosinophil counters the action of many mast cell mediators by
secreting degradative enzymes, including histaminase and aryl sulphatase,
which destroy histamine and leukotrienes respectively. In helminthic
infestation, major basic protein and eosinophil cationic protein released from
the eosinophil are probably beneficial in promoting elimination of the parasites. An elevated eosinophil count is typical in allergic reactions and in parasitic infestations, which support both these putative functions. Eosinophils chemotactically aggregate in large numbers to sites where antigen-antibody complexes are found and where the host is attempting to deal with foreign bodies. They're found easily in the connective tissue below a wheal, the raised "bump" that results at the site of an insect bite. The strong immune response such a bite elicits brings them in, and a section taken through a wheal will have thousands of eosinophils evident. Here's an example of such an eosinophilic exudate. Note the very bright and obvious staining of these cells, as well as the nuclear configuration. In an H&E stain, these could easily be mistaken for mast cells at this low magnification, but such huge aggregations are never seen in normal situations. Finding eosinophils in circulating blood is, frankly, a matter of dumb luck. They aren't anywhere near as numerous as neutrophils (they make up about 0.5-3.0 % of all white blood cells, depending on the species) so spotting one is usually an accident.
Granulocytes: Basophils
Basophils are by far the rarest of the granulocytes, and are believed to
be absent from cats, rats, and mice. In fact, in all species the basophil the rarest of all of the formed elements. Once seen, they're hard to mistake for any other cell type: in smears they have a deep purple, wine colored look to them, and the granules appear to be on the "outside" of the cell. They aren't, of course, like any granules they're intracellular. But the large granules in this rather small cell protrude considerably and stretch the plasma membrane. It sometimes looks as if the basophil had been coated with something and "rolled" in granules. The density of the staining is usually such as to obscure the nucleus, but when it's visible it usually is S-shaped.

 The true nature of basophils is still debated, but there's a general agreement that they're the circulating form of the common mast cell seen in the loose collagenous CT.
The true nature of basophils is still debated, but there's a general agreement that they're the circulating form of the common mast cell seen in the loose collagenous CT.
One argument supporting this assertion is the indisputable fact that basophils contain granules having the same materials in them as mast cells. The rarity of the basophil in circulating blood could easily be due to a very short "transit time" between leaving the bone marrow and taking up residence in the CT.
There's morphological evidence for the assumption. They're about the same size as
neutrophils, and the nucleus is usually bi-lobed of "S" shaped. The cytoplasmic granules are
coarse, and strongly basophilic. In a good preparation they stain a deep purple
color, often so strongly as to obscure the nucleus. Often basophil granules
will discharge their contents, or the materials in them may be leached out in preparation, all of which are characteristics of the mast cell in vitro and in vivo. Furthermore, the granules are known to contain histamine and heparin, both of which are found in
mast cells in connective tissues. In some animals the mast cells contain serotonin, a known neurotransmitter. Mast cells respond to some neurotransmitters by "degranulating" and releasing histamine: these materials affect basophils and cause their degranulation as well.
Don't expect to see a basophil in circulation: it would almost require Divine Intervention for this to happen. I won't say that nobody has ever done so, but in nearly 30 years of looking through microscopes, I can't say I've ever encountered one in a tissue section. Stick to blood smears for this cell type.
Agranulocytes: Lymphocytes
The lymphocyte is typically the most numerous and most important of the
agranulocytes. Lymphocytes come in several sizes, those in the blood being
characterized as "small" (5 to 10 microns) or "medium" types (10 to 18
microns). The "large" size is usually found outside the circulation, mostly in
lymphatic organs. Lymphocytes can be identified in smears by their very high ratio
of nucleus to cytoplasm. The nucleus is very densely stained, and usually round
(although it may have a very slight indentation). The cytoplasm is a thin band
to one side of the nucleus, and is stained a dusky blue color.

 The lone lymphocyte in the smear at right, adrift among vast numbers of erythrocytes, is pretty typical of its kind. Lymphocytes in circulation are small cells. This one is certainly no larger than the erythrocytes, and sometimes they're even a bit smaller. The extremely dense nucleus fills virtually the entire volume of the cell, and there is only a thin and tenuous rim of cytoplasm around it.
The lone lymphocyte in the smear at right, adrift among vast numbers of erythrocytes, is pretty typical of its kind. Lymphocytes in circulation are small cells. This one is certainly no larger than the erythrocytes, and sometimes they're even a bit smaller. The extremely dense nucleus fills virtually the entire volume of the cell, and there is only a thin and tenuous rim of cytoplasm around it.
Lymphocytes are found in many states in the CT, but in the circulation they're usually quiescent like this one. Once a lymphocyte emerges into the CT, and it's confronted with the "enemy" in the form of the specific antigen to which it responds, it will undergo transformation (at the site of the antigen or in a germinal center). If it's one of the B lymphocytes, it will become a plasma cell.
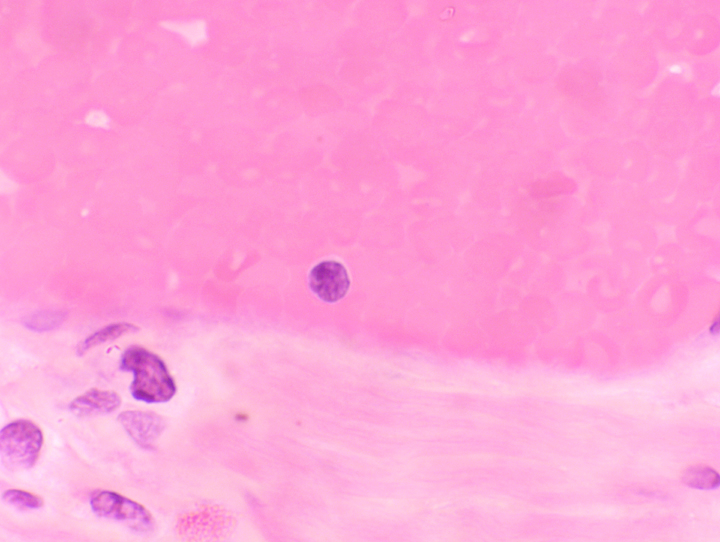 Lymphocytes are frequently seen wandering through the interstices of epithelial sheets, and scattered aggregations of them can be encountered in almost any organ. There are actually two functional types of lymphocytes, one responsible for cell
mediated immune reactions (T-cells) and the other for humoral immunity (B-cells)
but these can be told apart only by serological tests. They are morphologically
indistinguishable.
Lymphocytes are frequently seen wandering through the interstices of epithelial sheets, and scattered aggregations of them can be encountered in almost any organ. There are actually two functional types of lymphocytes, one responsible for cell
mediated immune reactions (T-cells) and the other for humoral immunity (B-cells)
but these can be told apart only by serological tests. They are morphologically
indistinguishable.
Because they're so numerous, it's usually easy to find lymphocytes in circulating blood.
Here's one, in a small vein. Identification is simplified by the round and very heterochromatic nucleus and the almost completely invisible cytoplasm. Although in stains the cytoplasm is often visible as a thin rim, in circulation it's typically lost in the background staining.
The sea of pink in this image represents large numbers of erythrocytes. For the most part they're not individually discernible, thanks in part to being tightly packed, and in part to the uniformity of their staining reaction. The lymphocyte is highlighted under these conditions and spotting it is simplified.
Agranulocytes: Monocytes
Monocytes are the second type of agranulocyte, and the largest of the
circulating formed elements. A healthy monocyte will run anywhere from 16 to 25
microns in diameter, much larger than any other formed element. The nucleus
varies in form, and may be kidney shaped, bean shaped, or (classically)
horseshoe shaped, with a deep indentation.

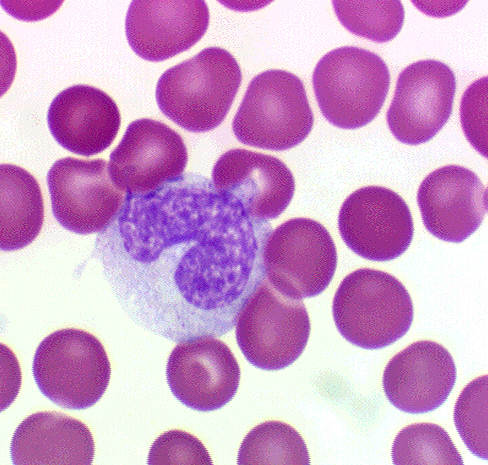 The nuclear material of monocytes doesn't stain as deeply as that of
lymphocytes, and chromatin strands can be made out (at least one author has
described it—in a remarkably colorful phrase—as a "spaghetti and meatball"
nuclear pattern). The ratio of nuclear material to cytoplasm is also less than
in the lymphocyte. The cytoplasm of the monocyte will stain a blue-gray color
and is said to have a "ground glass" appearance due to the presence of fine
granules in it.
The nuclear material of monocytes doesn't stain as deeply as that of
lymphocytes, and chromatin strands can be made out (at least one author has
described it—in a remarkably colorful phrase—as a "spaghetti and meatball"
nuclear pattern). The ratio of nuclear material to cytoplasm is also less than
in the lymphocyte. The cytoplasm of the monocyte will stain a blue-gray color
and is said to have a "ground glass" appearance due to the presence of fine
granules in it.
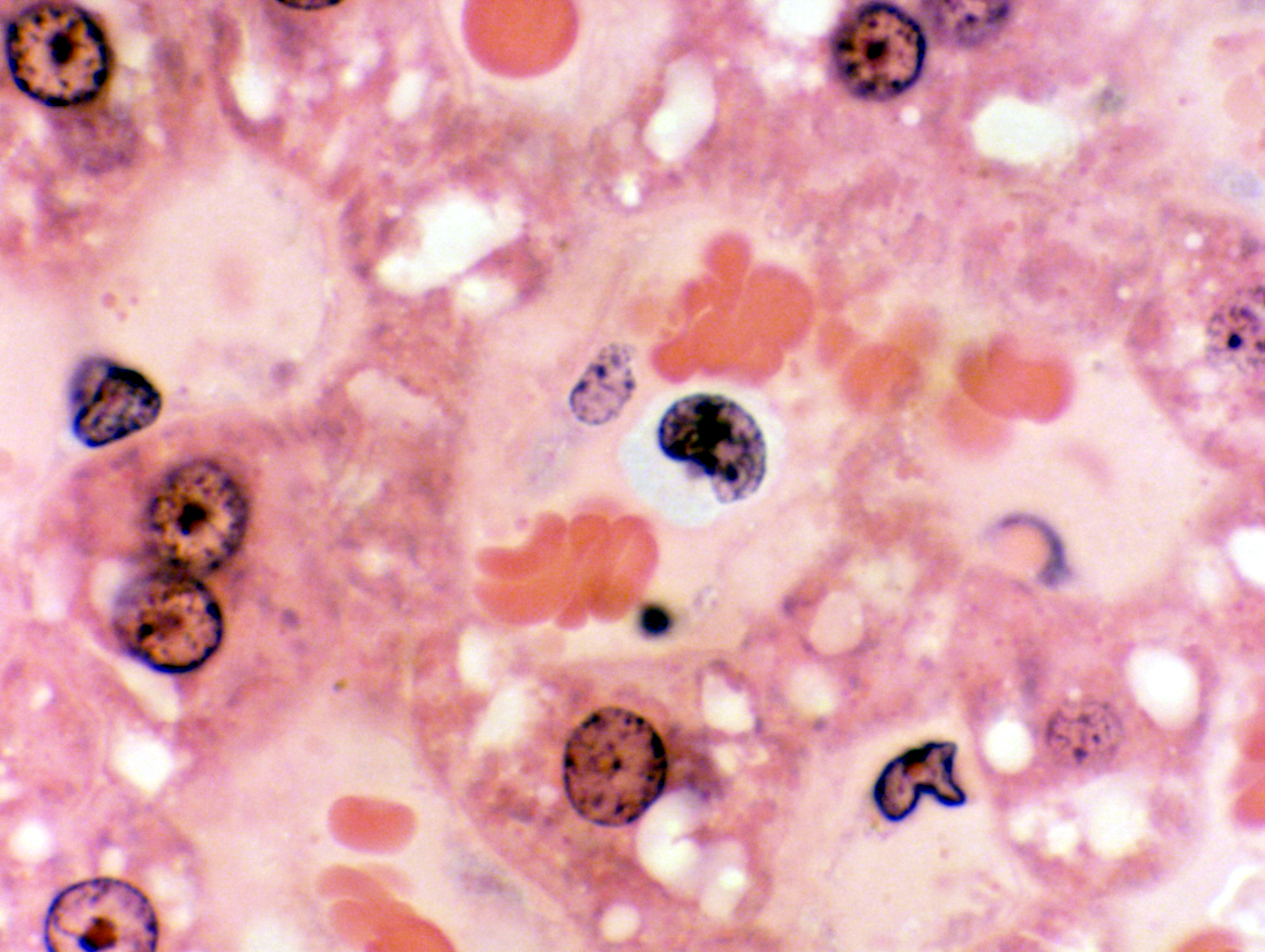 Monocytes aren't common in circulating blood, representing perhaps 5% to 8% of all leukocytes, but you do run across one now and then. When you do spot one, it's pretty obvious what it is, based on the size alone. In addition, the greater amount of cytoplasm and the deeply indented nucleus (depending on the geometry of the section this may or may not be visible) are confirmation. In the image shown here, the monocyte is by far the largest cell in the field. The indentation of the nucleus isn't pronounced, thanks to the orientation of the cell with respect to the plane of section, but the very significant amount of cytoplasm visible is: no lymphocyte would have as much. the slate-blue color of the cytoplasm is another clue. This cell is easily three times the size of the erythrocytes that are located near it. This cell was spotted on a slide of the liver, in one of the blood sinusoids; a good place to go hunting for circulating leukocytes.
Monocytes aren't common in circulating blood, representing perhaps 5% to 8% of all leukocytes, but you do run across one now and then. When you do spot one, it's pretty obvious what it is, based on the size alone. In addition, the greater amount of cytoplasm and the deeply indented nucleus (depending on the geometry of the section this may or may not be visible) are confirmation. In the image shown here, the monocyte is by far the largest cell in the field. The indentation of the nucleus isn't pronounced, thanks to the orientation of the cell with respect to the plane of section, but the very significant amount of cytoplasm visible is: no lymphocyte would have as much. the slate-blue color of the cytoplasm is another clue. This cell is easily three times the size of the erythrocytes that are located near it. This cell was spotted on a slide of the liver, in one of the blood sinusoids; a good place to go hunting for circulating leukocytes.
Be aware that there's a bit of a size overlap between large lymphocytes and small monocytes. Distinguishing which is which can take some practice and a sharp eye for details like nuclear configuration and colors. The best rule to use is that if you aren't sure it's a monocyte....it probably isn't.
Non-Mammalian Blood
The blood of non-mammals is somewhat different: the most conspicuous difference
is that all non-mammals have nucleated erythrocytes. The presence
of nuclei in significant numbers of mammalian erythrocytes is a sign of pathology, but in non-mammals
it's normal. In this case, the Greek root kytos (for "cell") has real meaning, because in all non-mammals, the erythrocyte of the circulation is a true cell, complete with nucleus. As in mammals, its function is to carry oxygen to the tissues.
You'll most commonly encounter non-mammalian blood in the form of avian blood.
Slide 642, of the
avian tongue, contains an area of hemorrhage which provides an opportunity to
see typical erythrocytes. Avian erythrocytes (and non-mammalian ones in general) are oval in shape, and
have a distinct centrally located nucleus. Although size varies somewhat with
sex and breed, avian erythrocytes are typically about 6.0-8.0 µ wide and
9.0-12 µ long. The nucleus is strongly basophilic and the cytoplasm
eosinophilic. Frequently you will be able to make out a clear area around the nucleus; this is
an enlarged perinuclear space. The space is real—it's present in all nucleated
cells—but its enlargement to the point where it can be seen in a light
microscope is probably a preparation artifact. The image below at left is from a chicken; the one at right is from a frog. The similarities are obvious.


The thrombocyte, also seen in this field from slide 642, is the non-mammalian equivalent of the platelet, and it has the same function of on-the-spot repair of breaks in the vascular system. In some clinical contexts the word "thrombocyte" is used interchangeably with "platelet" in mammals, which strictly speaking isn't correct usage. In birds, however, it is: the thromobocyte is a true cell, complete with nucleus and other organelles.
Thrombocytes are smaller than erythrocyte and in
smears have a faintly bluish tinge to their cytoplasm. One immediately
recognizable feature in good preparations is a small eosinophilic vacuole
located at one end of the nucleus. It appears as an orange dot in most
preparations. This perinuclear "dot" is part of the Golgi apparatus.
Home Page
Lab Exercise List