SLIDES FOR THIS LAB: 1203, 1210, 1205, VM58, VM62, 1-8, 85
PRE-RECORDED LECTURE FOR THIS EXERCISE
The oral cavity is the first portion of the digestive tract, and consists of the mouth and the structures which are enclosed within it. The mouth isn't merely a hole in the face. It's a complex structure with nutritional, respiratory, and communicative functions in all mammals and most other groups of animals.
Additionally, it serves as the principal means of manipulating (if you'll pardon the expression) the environment for those animals which don't have hands, such as dogs. A dog carries things in his mouth because he has no other way to do it. (If you have time someday, try to get a dog to bark when he has something in his mouth. This takes some work, but the noise he makes when trying to eat and communicate simultaneously is worth hearing.)
One can get a clue to the nature of an animal's diet by the type and shape of the teeth, which differ greatly in carnivores and herbivores.
Accessory structures which contribute to the mouth's function include the tongue, the teeth, and the salivary glands. The tongue and the teeth will be dealt with below: click here for a discussion of the structure of salivary glands.
The oral cavity in domestic animals is lined with stratified squamous epithelium. Depending on the nature of the animal's diet, this may or may not be keratinized. Those animals which eat a great deal of roughage, such as ruminants and horses, usually have a very heavily keratinized oral cavity, while those which eat softer food show less of it. There is a considerable amount of variation in this rule, both across species lines and among individuals in a species.
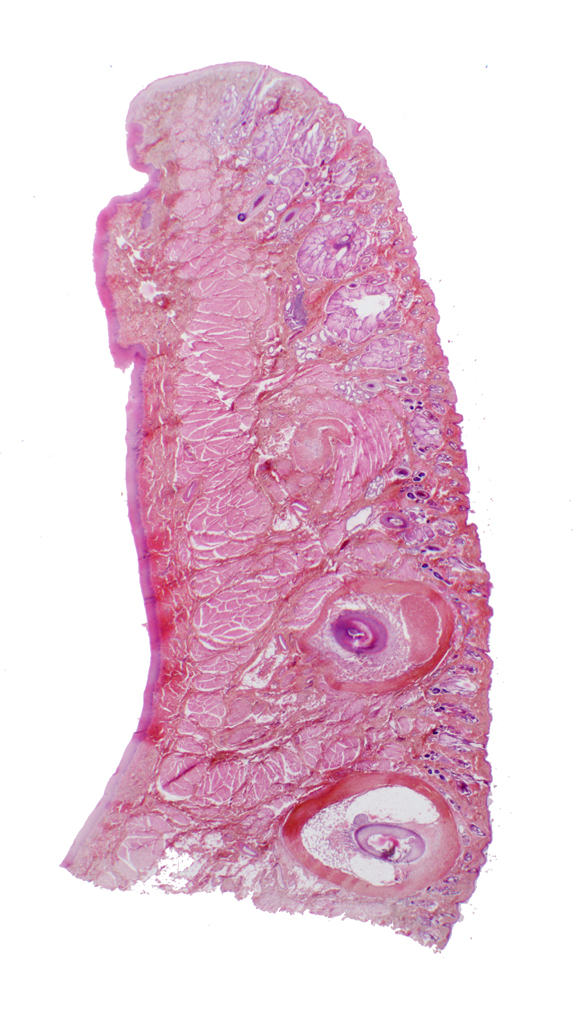
This is a section through the wall of the oral cavity, i.e., a cheek or lip. It's cut vertically, and you are looking into it from the side.
The oral side has a lining of stratified squamous epithelium. Since this specimen is from a primate, it's not keratinized. The oral cavity side totally lacks any of the adnexal structures associated with skin: no hairs, sebaceous or sweat glands, etc. The skin side has these things, and as external integument, it's always keratinized to some degree. The sinus hairs are the very large ones associated with tactile sensation through the vibrissae.
The center of this specimen contains some of the buccal muscles, strands of skeletal muscle that are part of the muscle sheet underlying the skin. Here it's acting as the "tunica muscularis" for this part of the digestive tract. Many rodents, such as the Golden Hamster (Mesocricetus auratus) have cheek pouches for temporary storage of food. The cheek pouch muscles are very well developed. The hamster fills the pouch and then can contract the buccal muscle to dump the contents whenever he wants.
The place of continuity between the "dry" integument and the "wet" oral mucosa is located at the top of this section. This sort of transition point is a mucocutaneous junction. Mucocutaneous junctions exist in several other sites: the anus, the nostrils, the vulva and the urethra.
Slide 1210 is a section of a tongue. The tongue in mammals is an
extremely muscular o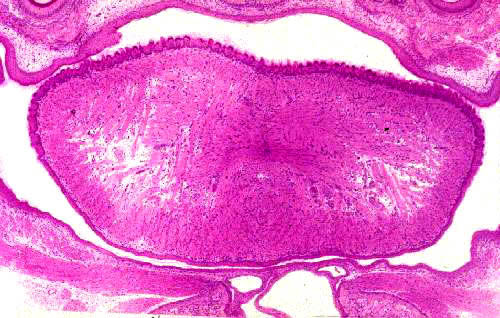 rgan within whose substance there are a number of smaller
salivary glands, and any number of motor and sensory nerve fibers. The
bulk of the tongue is skeletal muscle, arranged in three layers, all at right
angles to each other. This provides for an amazing degree of flexibility
and is vital to vocalization. There may be a considerable amount of
adipose tissue present as well. The orientation of the muscle layers
gives the tongue a distinctive appearance in LM sections that isn't easily
mistaken for anything else.
rgan within whose substance there are a number of smaller
salivary glands, and any number of motor and sensory nerve fibers. The
bulk of the tongue is skeletal muscle, arranged in three layers, all at right
angles to each other. This provides for an amazing degree of flexibility
and is vital to vocalization. There may be a considerable amount of
adipose tissue present as well. The orientation of the muscle layers
gives the tongue a distinctive appearance in LM sections that isn't easily
mistaken for anything else.
 The dorsal surface of the tongue is covered with specialized lingual papillae (see below), special regions of the stratified squamous epithelium that covers the entire organ. Not much detail can be see in this low power image, but a closer look will reveal the way the muscle bundles are laid out. In this higher magnification view, the muscle bundles and their arrangement can be seen clearly. This organization of muscle bundles is one of the identifying features of the tongue. One set runs parallel to the screen from upper left to lower right; a second set runs at a right angle to these from lower left to upper right. The third group is cut in cross section, as it runs in and out of the plane of the screen; it's therefore at right angles to both the other two. The bundles are thus set into X, Y, and Z axes. The muscle bundles play off against one another, and by controlling the tension it's possible for the tongue to assume many shapes.
The dorsal surface of the tongue is covered with specialized lingual papillae (see below), special regions of the stratified squamous epithelium that covers the entire organ. Not much detail can be see in this low power image, but a closer look will reveal the way the muscle bundles are laid out. In this higher magnification view, the muscle bundles and their arrangement can be seen clearly. This organization of muscle bundles is one of the identifying features of the tongue. One set runs parallel to the screen from upper left to lower right; a second set runs at a right angle to these from lower left to upper right. The third group is cut in cross section, as it runs in and out of the plane of the screen; it's therefore at right angles to both the other two. The bundles are thus set into X, Y, and Z axes. The muscle bundles play off against one another, and by controlling the tension it's possible for the tongue to assume many shapes. 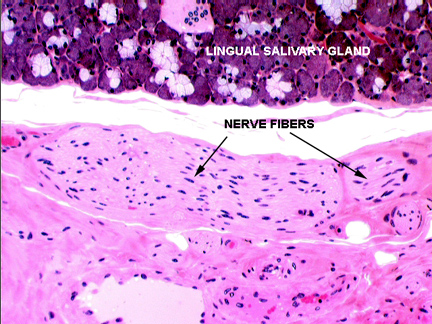

On the tongue's dorsal surface slide 1205 shows numerous filiform papillae. These are slender conical structures, which project upwards forming a velvety covering on the tongue. The filiform papilla is one of 4 types of lingual papillae, and its function is purely mechanical; it has no sensory structures associated with it, as do the others. In some species (such as cats and cattle) filiform papillae are extremely large and rough, and the peculiar rasping effect of a cat's tongue is due to these papillae. It's also what a cat uses to clean her fur. The stiff papillae make effective bristles for removing debris. You can also see some of these on slide 115.
The filiform papillae are by far the most common type, and sometimes are "swallow tailed" in shape. They're raked backwards. The "spines" on a cat's tongue are a nice example of this type of papilla, and they're what gives the tongue its rough feel and velvety appearance. They have no sensory function. In large animals, such as cows, the filiform papillae can be very large, up to perhaps 5.0 mm long. When they get this big, they sometimes have a "core" of CT inside. But in smaller animals they are pretty much entirely epithelial in nature.
Other Lingual Papillae
The remaining types of lingual papillae are sensory in function. They include: the fungiform papillae, which are less common than the filiforms; the vallate (or circumvallate) the largest and most prominent; and the foliate papillae, which are most easily seen in the Order Lagomorpha (rabbits, hares, and conies).
Vallate Papillae

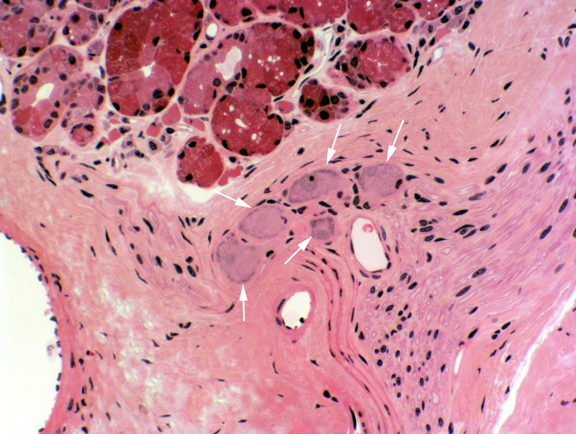
A vallate papilla is present on slide 1210. This type of papilla is set into a deep pocket in the tongue's surface, and anchored at the bottom by a short broad stalk. It doesn't protrude above the general level of the surface by much, but it's surrounded by a deep "moat" into which some of the lingual salivary glands secrete. Vallate papillae are easily visible with the naked eye in most animals. They're paired and located near the back of the tongue. They're the least numerous type. This very large structure has a core of connective tissue (CT) and nerve fibers are often seen in it, serving the taste buds.
Taste buds (TB) are present on the sides of the papilla, and also frequently on the side of the moat. The small lingual salivary glands (SG) that empty into the moat keep the taste buds ready for re-use. There are two types of these lingual salivary glands: serous and mucous in nature. The secretions of these glands are wash the chemoreceptor sites of the taste buds free of molecules, so that new sensations can be detected. These aren't the only salivary glands in the tongue, by the way; there are small ones scattered throughout the muscle bundles.
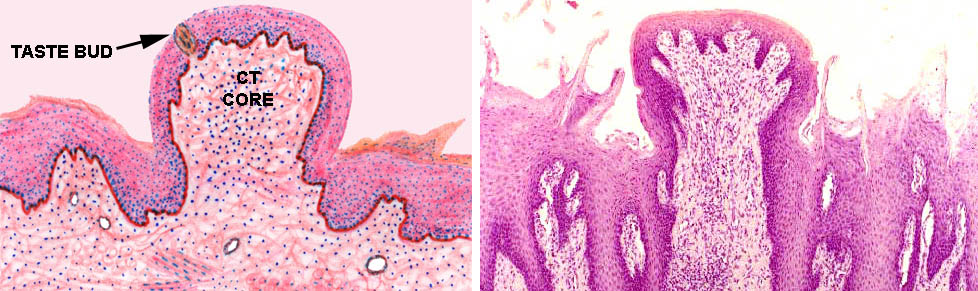
Vallate papillae usually show taste buds, seen as well defined, lightly staining areas which enclose several banana shaped sensory transducer cells. You should see some on the undersides of the papilla proper, and you may also see some on the tongue side of the "moat." Taste buds are discrete and well encapsulated by delicate CT, which can be demonstrated with special stains.
The sensory cells of the buds are located below the level of the epithelium, and they communicate with the outside via a so-called taste pore, a break in the epithelium. Through this pore project the microvilli or "taste hairs" on the tops of the sensory cells. The taste bud is yet another example of a peripheral transducer structure whose function is to convert some physical phenomenon into a nervous signal. In this case the phenomenon is the binding of tastant molecules to the membrane of the transducer cells. Binding causes charge changes on the cells that are detected by neural fibers wrapped around them.
Not a whole lot is understood about the phenomenon of taste. Traditionally there are said to be four basic taste sensations: sweet, salt, sour, and bitter, but recent research casts some doubt on this simplistic explanation. Specific regions of the tongue do have specific associations with these sensations: the data is incomplete, but it's been shown that generally the tip of the tongue has receptors for sweet and salty sensation, and sour and bitter receptors lie near the back. There are no morphological differences that correlate with functional ones, however, and you can't tell by looking at a taste bud what kind it is.
It's thought that the different taste sensations are located in and associated with specific cell types in the taste buds, but there are also indications that some cells may respond to more than one taste stimulation. Depending on where they're located, the taste buds may be innervated by fibers from cranial nerves VII (facial), IX (glossopharyngeal) or X (vagal). This is separate from the "general" sensation; that is, tactile perception is not connected through the same nerve circuits. A branch of cranial nerve V (trigeminal) handles this input.
Taste sensation is found in all vertebrates, and a similar chemoreception ability in most if not all invertebrates. The more or less universal distribution of chemoreception makes it clear that this sense has very high survival value.
The receptor cells, regardless of type, are more or less banana shaped, and at the apex of each is a small microvillus, called a "taste hair." (This is a misnomer because they aren't "hairs" in any real sense, but we're stuck with this archaic name.) The "taste hairs" collectively come to form a bundle that's exposed to the world via a small opening in the overlying epithelium, the taste pore. The "hairs" are the actual site of chemoreception; the plasma membrane that covers them contains receptor sites for the different sensations.
Integration of the chemoreception signals takes place in the central nervous system. The transducers respond, but the brain actually "tastes" by interpreting the responses. To a great extent, perception of tastes as "good" or "bad" is learned behavior. In some cultures, grilled meat is considered delectable, but in others, nauseating. Similarly, I have lived in places where rotten eggs are regarded as a delicacy, but my cultural perceptions prevented me from eating them. Many animals (humans included) have to learn what foods are edible, and one of the clues they use is taste perception.
Fungiform and Foliate Papillae
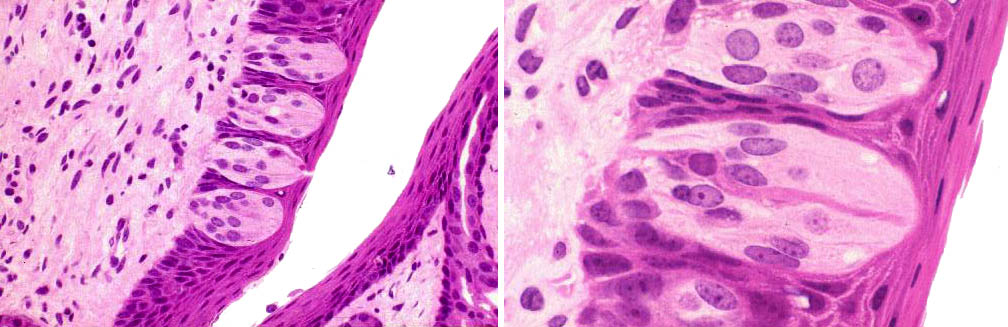
The fungiform papillae, as the name implies, are mushroom shaped structures. They rise above the general level of the filiform papillae, and they usually have taste buds. The foliate papilla is best seen in rabbits. The term means "leaflike" as in the leaves of a book, and these are seen in the demonstration as if you were viewing the top edge of a page of paper. They are set in the sides of the tongue, and you are looking at them from the dorsal surface. Both fungiform and foliate papillae carry taste buds.
The mushroom-shaped fungiform papilla is nicely illustrated here in diagram and in an actual specimen. Fungiform papillae are far less numerous than the filiform type, and scattered in among 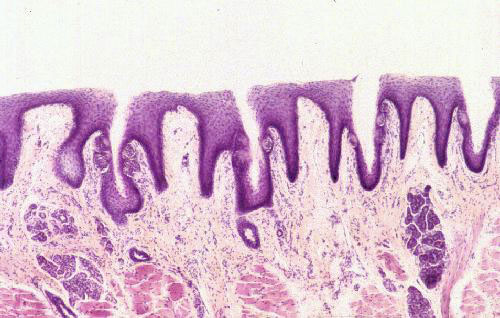 them. They're not nearly as large as the vallate papillae, but they are large enough to have a core of CT. The fungiform papillae stick up above the surrounding filiforms, and here you can see the relationship pretty well. A swallow-tailed filiform papilla flanks this fungiform one on either side.
them. They're not nearly as large as the vallate papillae, but they are large enough to have a core of CT. The fungiform papillae stick up above the surrounding filiforms, and here you can see the relationship pretty well. A swallow-tailed filiform papilla flanks this fungiform one on either side.
Here's a group of foliate papillae. The word "foliate" means "leaflike" and in this case the reference is to the "leaves" of a book. The orientation of this specimen is different from the ones seen before. You are viewing these papillae from the top; that is, the tongue has been cut parallel to its long axis, and exposed to view from the dorsal side. Thus, you're seeing the edges of the foliate papillae as if you'd seen the top edge of a book.
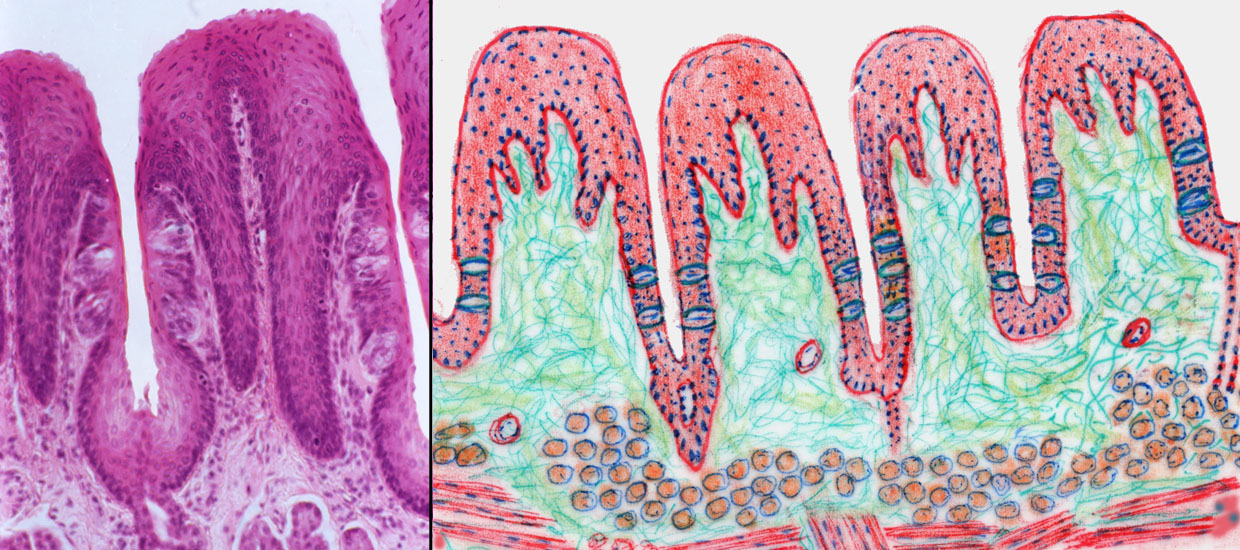 The papillae themselves are flat-topped and have deep clefts between them. The clefts are lined with epithelium, and there are taste buds embedded in them. This type of papilla is most commonly seen in members of the Order Lagomorpha, i.e, rabbits, hares, pikas, and conies. Similar papillae are found in the tongue of the Florida manatee!
The papillae themselves are flat-topped and have deep clefts between them. The clefts are lined with epithelium, and there are taste buds embedded in them. This type of papilla is most commonly seen in members of the Order Lagomorpha, i.e, rabbits, hares, pikas, and conies. Similar papillae are found in the tongue of the Florida manatee!
Sound teeth are one of the more important things in life, and an abscessed tooth is likely to make the whole world look black to the individual suffering from it, two-legged or four-legged. Yes, dogs do get dental caries, and they can suffer from them, though they're less likely to have them than humans are because of differences in diet and tooth shapes.
 We featherless bipeds, and most of our pets, have brachydont teeth,
like the one shown at left. This is the type found in carnivores and the "typical" tooth model. (Ruminants and
horses have a different type which won't be considered in this exercise.)
You will find a typical brachydont tooth on slide 85. The tooth on this slide
is developing in a socket of the jawbone. This bony alveolus is
part of the skeleton, but strictly speaking the tooth isn't.
We featherless bipeds, and most of our pets, have brachydont teeth,
like the one shown at left. This is the type found in carnivores and the "typical" tooth model. (Ruminants and
horses have a different type which won't be considered in this exercise.)
You will find a typical brachydont tooth on slide 85. The tooth on this slide
is developing in a socket of the jawbone. This bony alveolus is
part of the skeleton, but strictly speaking the tooth isn't.
There are three hard materials involved in tooth structure: enamel, dentin, and cementum. Enamel covers the outside of the tooth, and in the processing of a tooth for microscopic examination, it's usually lost. It's a thin, brittle coat that's easily chipped and almost entirely made of inorganic materials. Dentin comprises the bulk of the tooth's mass. It's softer than enamel, and underlies it. Cementum is a bone-like substance with embedded cementocytes, that covers the root of the tooth, but doesn't project above the gum line.
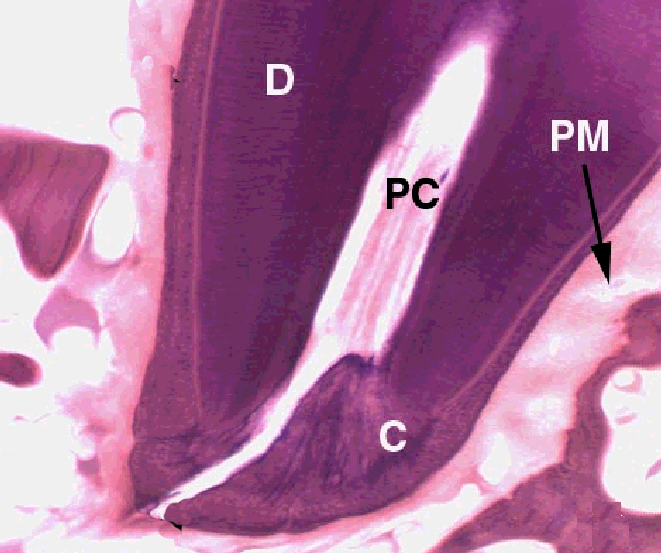 Brachydont teeth are hollow. There's a space inside, the pulp cavity, that contains living material in the form of cells, and a relatively undifferentiated primitive form of CT. The pulp cavity has blood vessels that run in and out to serve the needs of these tissues, and also nerve fibers.
Brachydont teeth are hollow. There's a space inside, the pulp cavity, that contains living material in the form of cells, and a relatively undifferentiated primitive form of CT. The pulp cavity has blood vessels that run in and out to serve the needs of these tissues, and also nerve fibers.
Since the tooth has to have a blood supply, and since only CT can make blood vessels, the periodontal ligament serves not only as the anchoring material, but as the conduit through which blood vessels and nerves go to and from the pulp cavity. Access to this space is via the root canal, visible in the image to the right as a channel through the hard material. When a dentist does a "root canal" on you, what's happening is the entire pulp cavity is being purged of its contents, and filled with restorative material. He has to remove the blood vessels and nerves, which is done by drilling out not only the pulp, but all the way down through the root canal, and then stuffing everything full. The presence of the nerve fibers is what makes this such an exhilarating experience.
Teeth are anchored into the bone of the jaw by a tough, collagenous periodontal ligament (sometimes called the periodontal membrane, PM above) that serves as the periosteum for the bony socket, and attaches firmly to the base of the tooth, where the cementum (C) is located. Try this: grab one of your teeth and push it, fairly hard: it will move, just a bit. That's the "give" in the ligament. Sometimes teeth get loosened by an impact; when this happens after a while the ligament will repair the damage and the tooth is firmly seated once more.
Tooth formation is a complicated process, and the tooth's hard structures are the result of cellular activity, but are in and of themselves non-cellular, dead material. Primitive connective tissue, or mesenchyme condenses in the future central region of the tooth, in what will become the pulp cavity. Inside this region a special population of cells, the odontoblasts begin the process of making dentin, by laying down a matrix material and then calcifying it. As they do so, they move backwards towards the center of the pulp cavity. The odontoblasts remain active throughout life, so the process of making dentin continues after eruption. As the animal ages the pulp cavity slowly diminishes in size.
On the outside surface of the tooth, a similar process takes place. A population of ameloblasts differentiates, as a row of nicely columnar cells facing the odontoblasts. Ameloblasts make enamel in a way that's comparable to the manufacture of dentin. They lay down an uncalcified matrix first, then harden it, backing away as the enamel layer is built up. In brachydont teeth, enamel is made only before eruption. Once the tooth erupts, the ameloblasts die. If enamel is damaged it can't be renewed.
You see here a typical forming tooth, in situ. An important thing to remember is that the hard material of the tooth—the enamel and the underlying dentin—are both formed extracellularly.
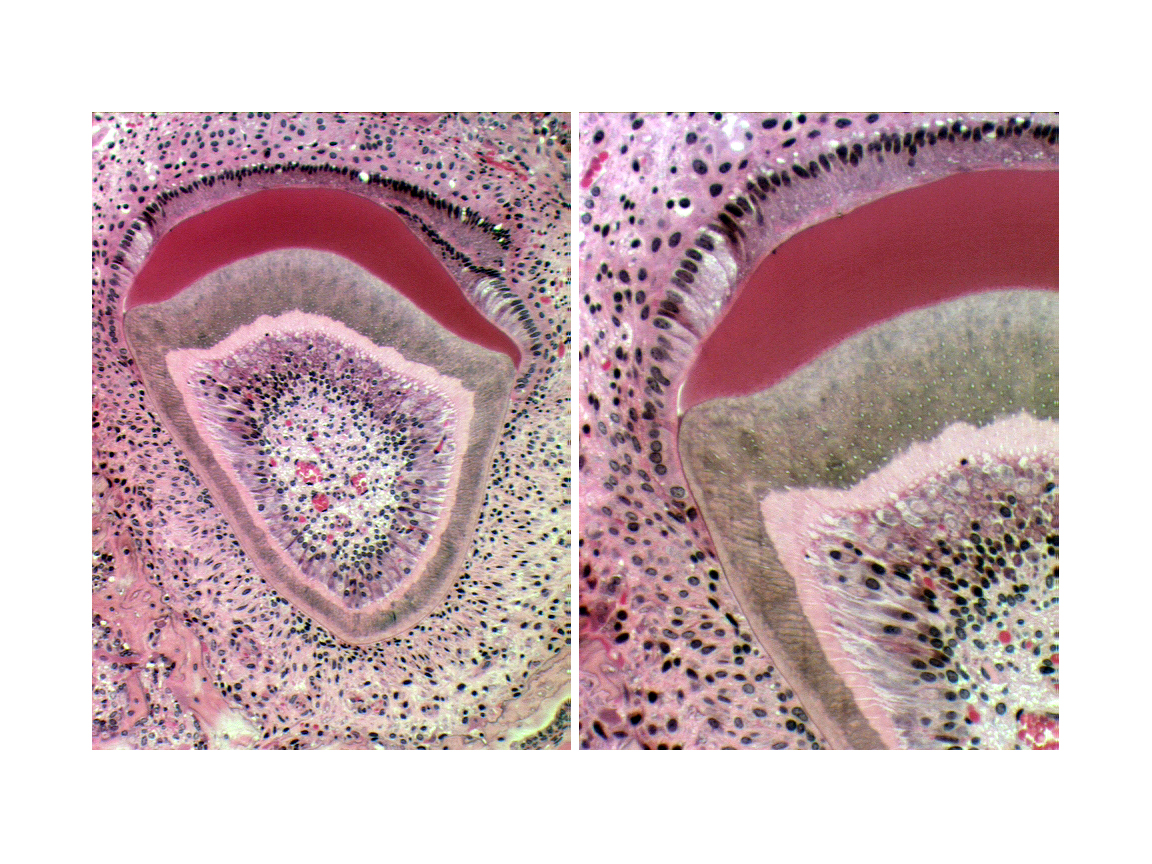
The ameloblasts are ectodermally derived, from the oral epithelium in the embryonic oral cavity. The embryonic origins of the odontoblasts, the cells which make the dentin, is a matter of some debate, but they are resident in the site of the future pulp cavity, which is generally held to be embryonic mesenchyme. Hence, they are probably derived from mesoderm. There is recent evidence to suggest, however, that they may be one of the many derivatives of the neural crest cells, and ectodermal as well.
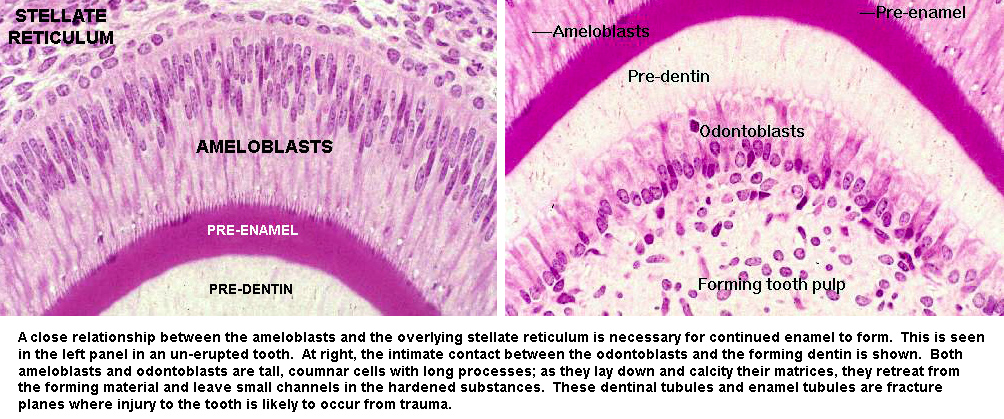
Odontoblasts and ameloblasts have some similarities, whatever their origins. Both cell types are tall and columnar in shape. They lay down precursor materials—pre-dentin and pre-enamel—and as they do so, "back away" from it. The precursor material is then calcified as a new layer of precursor is formed next to it. In the course of tooth formation, differential deposition of dentin and enamel creates the proper shape of the tooth. This specimen comes from a mouse fetus, and it's early enough in fetal development that no actual calcification of the enamel has yet occurred, hence it's really "pre-enamel." Dentin calcification hasn't yet taken place, so pre-dentin is visible.
The odontoblasts live in a nice protected environment and continue their activity throughout life. The walls of the pulp cavity are made of dentin, of course, lined by the active odontoblasts. The inductive stimulus the ameloblasts need comes from the CT overlying them as the tooth bud forms. 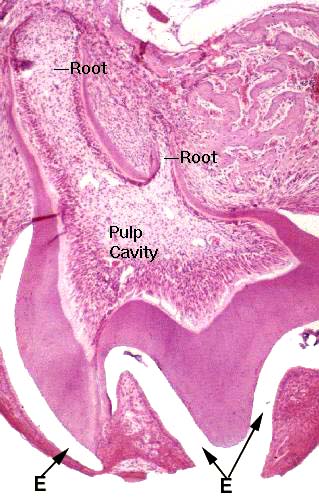 This material, the stellate reticulum, originates as part of the mesenchyme of the head. Once the tooth erupts through the gumline, the cells of the stellate reticulum die, the stimulus is lost, hence the ameloblasts die. Enamel can't be made anew* and injuries to it are permanent.
This material, the stellate reticulum, originates as part of the mesenchyme of the head. Once the tooth erupts through the gumline, the cells of the stellate reticulum die, the stimulus is lost, hence the ameloblasts die. Enamel can't be made anew* and injuries to it are permanent.
Enamel is hard but brittle. the retreat of the ameloblasts leaves small channels through its substance, which are weak points. An impact can cause a fracture along these planes of cleavage, and if it's bad enough it can expose the underlying dentin. Damage to the enamel can also occur through dietary exposure and bacterial infection. If the enamel is damaged and not repaired, the dentin in turn is subject to mechanical injury or erosion. Eventually, the dentin will be perforated and the pulp cavity exposed to infection. This will result in serious problems; in addition to almost certain loss of the tooth, a really bad infection can spread to the surrounding jaw bone.
The image at left is from an 18-day mouse fetus, once that was very close to term (21 days in mice). It's a molar. It hasn't yet erupted, but it's close to the gum, and had this mouse been born, shortly afterwards the tooth would have broken through. The enamel is gone but the space where it was (E) is marked. This is a processing artifact: teeth have to be decalcified before cutting. Dentin persists because it has much more organic material than mineralization, so something is left behind to take the stain.
The complex shape of this tooth indicates how exquisitely the process is coordinated. The ebb and flow of inductive signals in the formation of the hard materials actually sculpts the proper shape for the tooth in question, be it a molar, a canine, or an incisor. The root formation is similarly controlled by intercellular signaling that switches on and off as needed; and by selective timed losses of cell populations and sub-populations. How this process is directed is not understood except in the most general sense.
*In some animals, especially in rodents, a portion of the enamel making apparatus remains viable below the gumline, and in these new enamel is made continuously. For rodents this provides a fresh, sharp, chisel-like edge to the front of the incisors. That's why they gnaw continuously: to wear away the excess and keep the edge sharp. If they are stopped from chewing their teeth will continue to grow until they puncture the roof of the mouth.