Digestive System: Esophagus and Stomach
The greatest part...of man's trouble is caused by the belly, the gratification
of which is the life's work of the majority of mankind...and so much have our
customs degenerated that it is chiefly through his food that a man dies. This,
the most troublesome organ in the body, presses as does a creditor, making its
demands several times a day. It is for the belly's sake especially
that...luxury uses spices, voyages are made...and the bottom of the ocean is
explored. Nobody...is led to consider how base an organ it is by the foulness
of its completed work.
—Pliny the Elder (23-79 A.D.)
Natural History, Book XXVI, Paragraph 28
Objectives for this Exercise
SLIDES FOR THIS LAB: 43, VM67, VM74, VM68, 133
PRE-RECORDED LECTURE FOR THIS EXERCISE
Alimentary Tract
General Structure of Tubular Organs
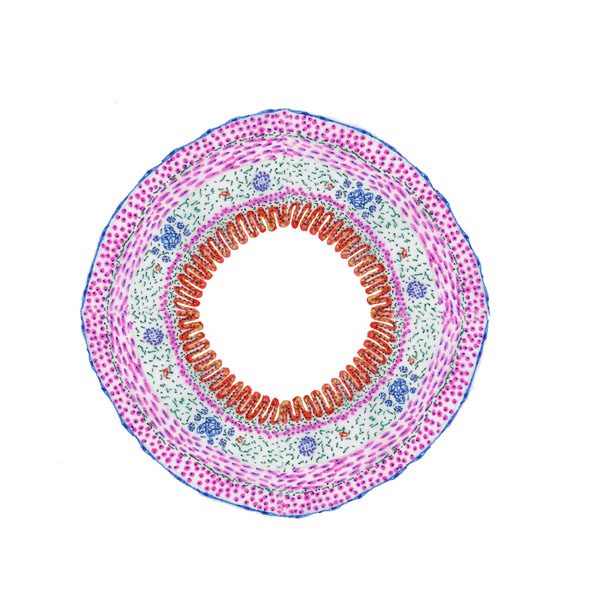 Once past the oral cavity, most of the digestive tract has a distinct structural
pattern that typifies tubular organs in general. Although there are
variations from place to place, especially in the nature of the lining epithelium and/or the presence of some structures, generally it includes four "tunics" or layers, a couple of which have subdivisions.
Once past the oral cavity, most of the digestive tract has a distinct structural
pattern that typifies tubular organs in general. Although there are
variations from place to place, especially in the nature of the lining epithelium and/or the presence of some structures, generally it includes four "tunics" or layers, a couple of which have subdivisions.
From the
innermost (i.e., closest to the lumen) to the outermost, these are the tunica
mucosa, the tunica submucosa, the tunica muscularis externa, and a tunica adventitia and/or tunica serosa. These are diagrammed at left.
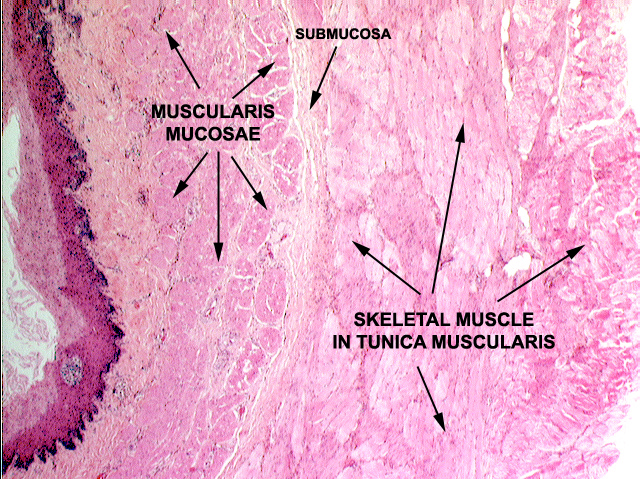
The tunica mucosa has three parts: the lining epithelium; the underlying lamina propria (a loose, cellular CT that supports the epithelium); and the muscularis mucosae, a layer of smooth muscle that separates the tunica mucosa from the tunica submucosa. The muscularis mucosae isn't always continuous, and it may send sprigs of muscle fibers up into the cores of villi where these are present. Its movements are controlled by a nerve plexus in the tunica submucosa (not labeled in the diagram). The tunica submucosa is another CT layer; it's more compact and robust than the lamina propria, and in it you will find blood vessels, nerve fibers, the usual complement of CT cells, and in some locations lymphatic structures or exocrine glands.
 Outboard of the submucosa, there are two (sometimes three) layers of muscle, the tunica muscularis externa, whose contractions are the propulsive mechanism for the food bolus or for the emptying of the organ. It has a large nervous complex between the layers, which will be discussed below.
Outboard of the submucosa, there are two (sometimes three) layers of muscle, the tunica muscularis externa, whose contractions are the propulsive mechanism for the food bolus or for the emptying of the organ. It has a large nervous complex between the layers, which will be discussed below.
If the organ is freely suspended in the body cavity (as, for example, is the greatest length of the intestine) it will be covered by a fold of peritoneum, called here the tunica serosa. This delicate covering is often lost in collecting samples of tissues. In places where the organ is attached firmly to surround structures it will have a third CT covering, a "tunica adventitia" as a means to anchor it in place.
We'll discuss the variations on this themes as we look at the different parts of the alimentary canal below.
The Esophagus
 The esophagus is a distensible cranial portion of the tract, formed as a
simple tube. At the top it receives a food bolus from the pharynx, and at the
bottom it discharges it into the first portion of the stomach. Start with slide
113; in the center of this slide is the esophagus of a rat, shown here at low magnification. Although few details are visible at this very low magnification, you can see that it is a muscular tube lined with epithelium. It's cut at about midlevel, with the dorsal side is at the top of the image.
The esophagus is a distensible cranial portion of the tract, formed as a
simple tube. At the top it receives a food bolus from the pharynx, and at the
bottom it discharges it into the first portion of the stomach. Start with slide
113; in the center of this slide is the esophagus of a rat, shown here at low magnification. Although few details are visible at this very low magnification, you can see that it is a muscular tube lined with epithelium. It's cut at about midlevel, with the dorsal side is at the top of the image.
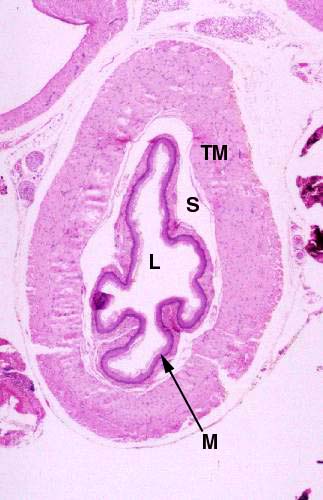
The lumen (L) is surrounded by the tunica mucosa (M); the lining of the esophagus in all species is stratified squamous epithelium, and in most animals it's keratinized, sometimes quite heavily. The tunica submucosa (S) is seen here as a space between the tunica mucosa and the muscularis externa (this is a shrinkage artifact of preparation). The tunica muscularis externa (TM) contains both skeletal and smooth muscle fibers. In some species (e.g. dogs and some ruminants) the entire tunica muscularis externa of the esophagus is composed of skeletal muscle; in birds it is entirely smooth muscle; and in most animals it makes a transition from skeletal to smooth muscle about half-way down (see below). These features can be used as landmarks to identify the organ and to determine where the section was taken.
The plicated nature of the tunica mucosa is obvious, even at this magnification. The esophagus has to be distensible to permit the swallowing of a fairly large bolus of food, and the mucosa is therefore "puckered" when the muscle of the tube hasn't been stretched. Although you can't see them here, there are also small glands in the submucosal layer that help to lubricate the food bolus in its passage. The contractions of the tunica muscularis push the food along its short journey to the stomach and these are controlled by an amazingly complex array of nerve fibers. The act of swallowing is a sort of high speed peristalsis, partially voluntary in nature, and very tightly controlled.
Esophageal Mucosa and Lamina Propria
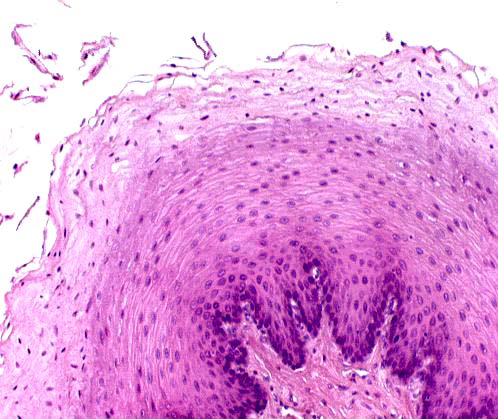 Here's the epithelial lining, at higher magnification. The epithelium here is clearly stratified squamous epithelium, and equally obviously, keratinized.
Here's the epithelial lining, at higher magnification. The epithelium here is clearly stratified squamous epithelium, and equally obviously, keratinized.
Stratified squamous epithelium is well suited to this particular location. The level of abrasion is high, especially in those species which eat a diet heavy in roughage. The proliferative nature of stratified squamous epithelium enables it to replace the surface that's constantly being worn away.
Some textbooks will tell you that "wet" keratinized stratified squamous epithelium doesn't develop a stratum granulosum, as the "dry" keratinized epithelium of the skin. This isn't true. Although this specimen is clearly from a "wet" location, the esophageal mucosa, the epithelium has a stratum granulosum.
One more clue to recognition of the esophagus is the presence of glands in the submucosa. These are referred to as esophageal glands proper, a term that implies they're a specific part of this organ, and are distinct from glands in the mucosal layer. Esophageal glands proper are outside the muscularis mucosae, and hence by definition in the submucosa.
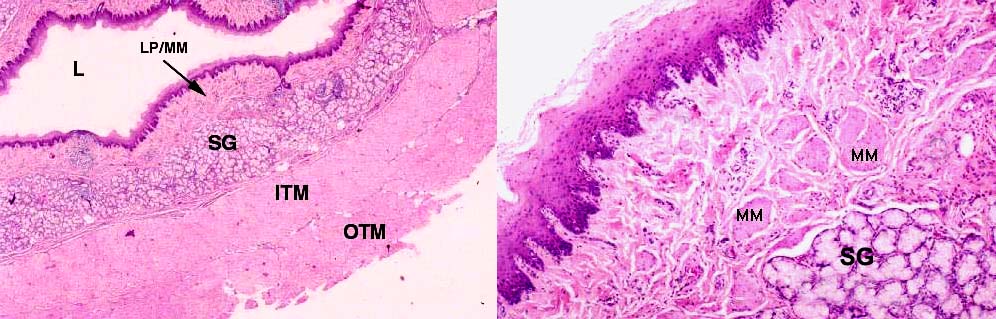
This image shows some of the submucosal glands, or esophageal glands proper (SG). Note the location of these glands with relation to the lumen and other structures. The muscularis mucosae (MM) in the esophagus is discontinuous (see below) but the esophageal glands proper are always outside the muscle bundles, hence they're in the submucosa. This is one of only two locations in the mammalian digestive tract where true glands are in the submucosa. The association of submucosal glands and a lining of stratified squamous epithelium is unique to the esophagus. The tunica muscularis is clearly defined as two layers, and inner (ITM) and outer (OTM) at right angles to each other.
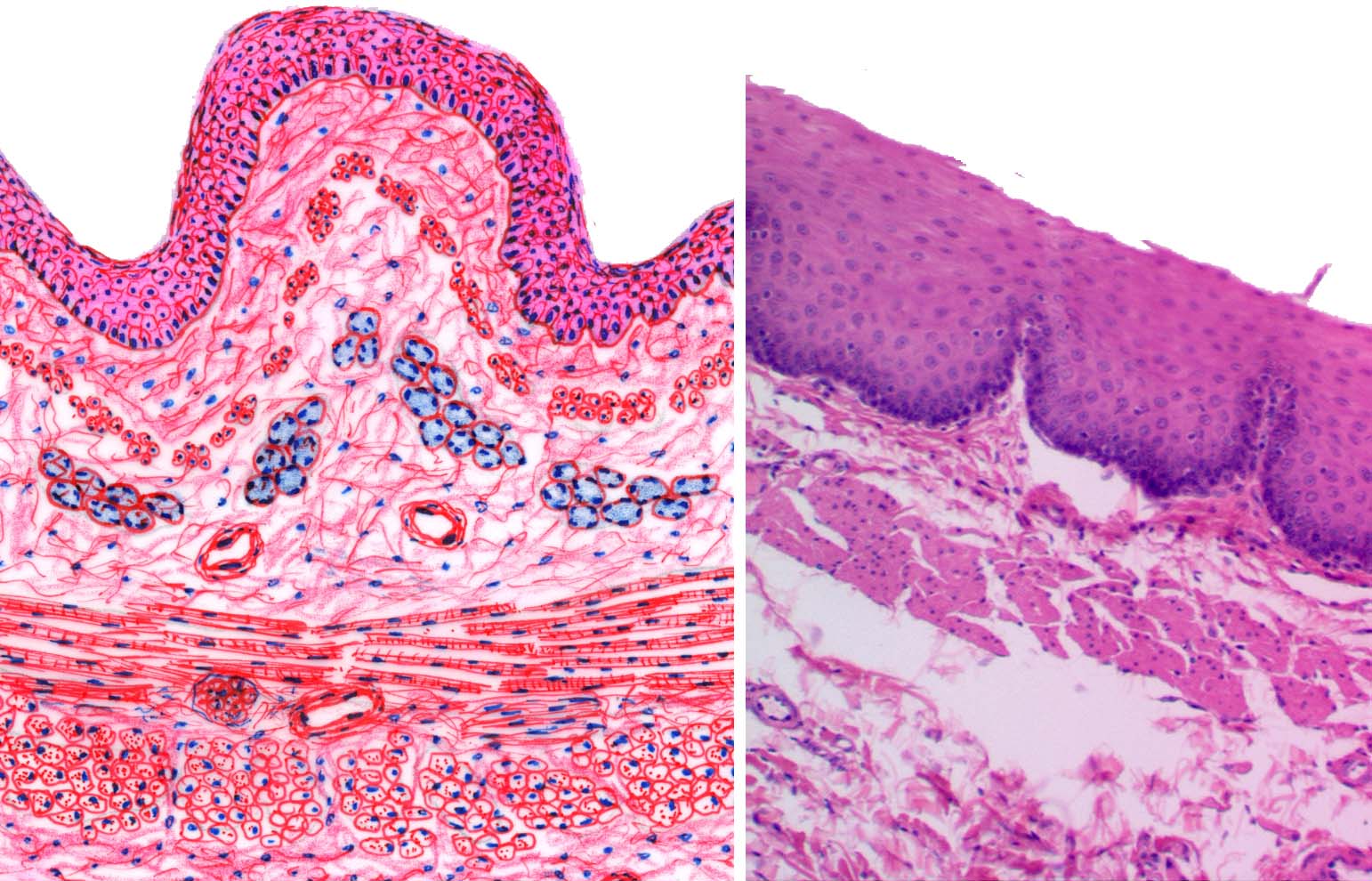
The muscularis mucosae of the esophagus is composed of smooth muscle fibers, running parallel to the long axis of the organ. In animals—unlike humans—the esophageal muscularis mucosae is discontinuous. The muscle is gathered into bundles that, when the organ cut in cross section look like "islands" of smooth muscle. The sketch above and the actual section can be compared to see this. The separation of the lamina propria from the submucosa is not so complete as it is in other regions of the alimentary canal, but nevertheless the muscularis mucosae is present and defines the two layers.
In the dog and the pig, the muscularis mucosae is absent in the cervical part of the esophagus, but becomes evident in the distal portions. On slide 43 you may see a few scattered bits of smooth muscle representing a muscularis mucosae in the cervical region. Near the stomach, the muscularis mucosae may become complete or nearly. In horses, cattle, and cats the cervical esophageal muscularis mucosae is it's discontinuous but present as scattered muscle bundles.
Esophageal Tunica Muscularis
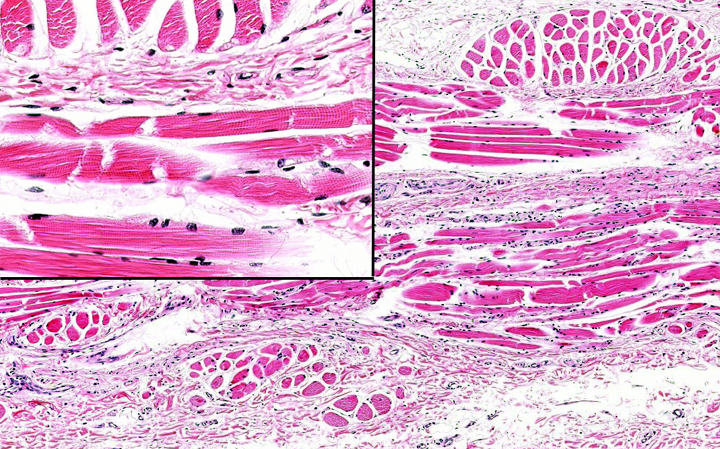
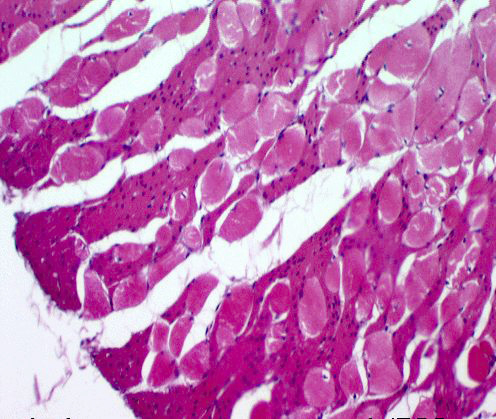
In the upper portion of the esophagus in most, if not all, mammals, the tunica
muscularis is composed of skeletal muscle arranged in two layers: inner
circular and outer longitudinal. You can see this in the image above. This is from the upper (cranial) esophagus and shows only the tunica muscularis. The inset shows a higher magnification of the muscle: striations are clearly visible in it, so it's skeletal, not smooth, muscle.
In most species, skeletal muscle is replaced
by smooth muscle as one descends deeper into the organ; the tunica muscularis
near the junction with the stomach is wholly smooth muscle in most animals. The mixed nature of the tunica muscularis in the esophagus is seen below: in this transition zone in a primate esophagus, where both type of fibers exist, there's a very large size disparity between them.
Dogs and ruminants are exceptions to this statement. In these animals, the
tunica muscularis is entirely skeletal muscle for the entire length of the
esophagus, which facilitates vomiting. In ruminants, regurgitation of swallowed
food is normal, and the "cud" that these animals chew is material
they have previously swallowed, then brought up again for reconsideration.
Although in dogs vomiting is technically not a "normal" process, there
are sound reasons for them to do it. Many wild dogs swallow small animals like
mice whole (see above). In the case of a bitch weaning pups, the only way to
get the food back to them is to gulp it down and barf it back up outside the
den, so the pups can have a nice hot meal. In domesticated strains the ability
to vomit voluntarily hasn't been lost.
Stomach
The stomach is a muscular bag whose principal function in most groups is
acidification and maceration of the food to the liquid state, and temporary
storage until it's passed to the intestines. Although nutrient absorption isn't a primary function of the stomach in
most species, it is certainly possible for material to be absorbed, and in some
groups gastric absorption provides a significant amount of energy intake. (In the ruminants the forestomach acts as a "fermentation vat" for the cellulose rich materials these animals
eat, and fatty acids produced there are absorbed across its mucosa.)
Esophageal-Stomach Junction
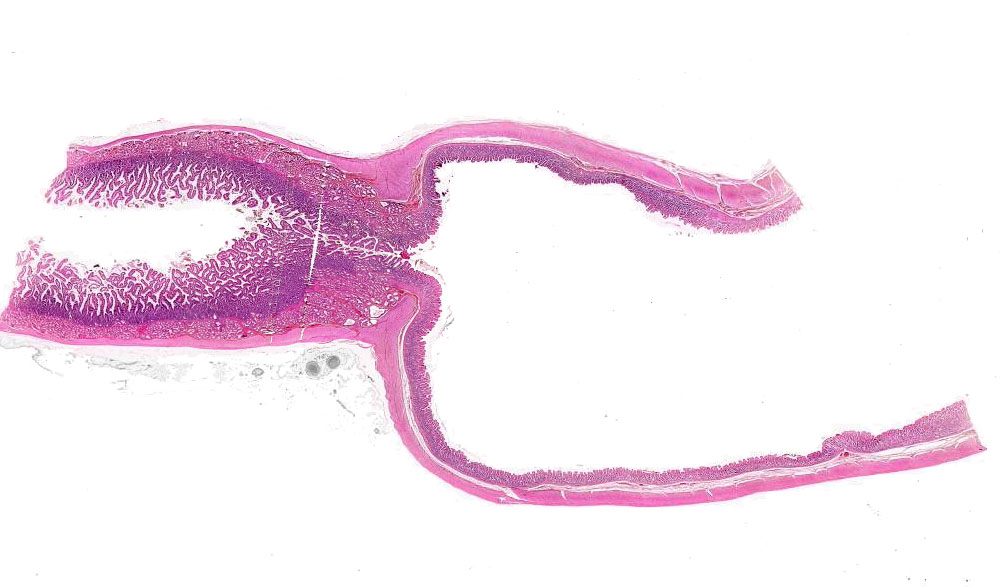
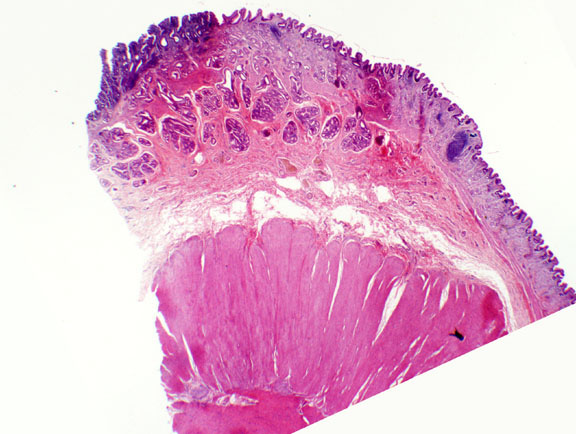
Slide 133 shows the esophageal-stomach junction. You can easily
differentiate between the two organs. There is stratified squamous epithelium
on the esophageal side and simple columnar epithelium on the stomach side. Note
the absence of goblet cells in the stomach.
In some species this abrupt change is located quite a bit deeper than it is in
others. The transition point between the two is what the gross anatomist calls
the margo plicatus inside the anatomic stomach. Horses are an example of
a species in which the transition is fairly deep, and so are rats.
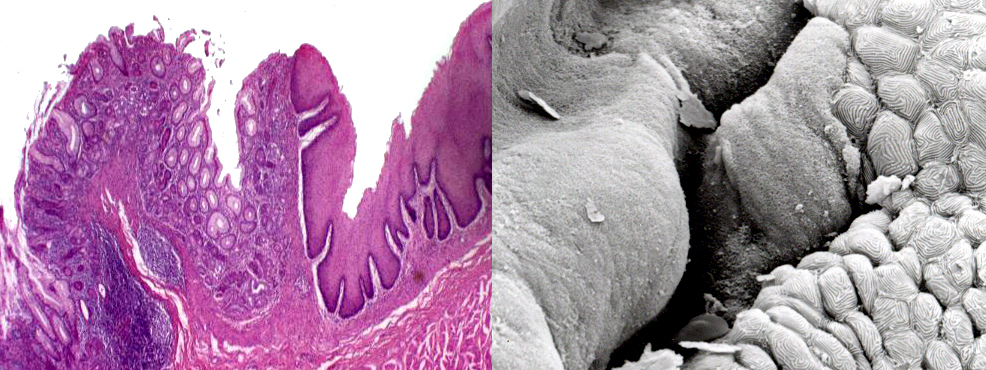
The images above are at the esophageal-stomach junction. On the right side of both the light micrograph and the scanning electron microscope image you see the stratified squamous epithelium of the esophagus which terminates abruptly at the point of entry into the stomach. The lining of the stomach is a simple columnar type without goblet cells. The stratified squamous cells of the esophagus have ridges that are visible in the SEM image, but the velvety appearance of the stomach portion is due to the millions of individual microvilli of its lining cells; the microvilli aren't individually resolvable at this level. Arrows demarcate the very abrupt line that separates the two types of lining epithelia.
There is no particularly thickened region of the tunica muscularis at this location. Although you will hear the term "esophageal sphincter" there isn't much in the way of a true anatomical sphincter at the entrance to the stomach, as there is at the pyloric opening (see below). The "esophageal sphincter" is physiological: intra-esophageal pressures are generally higher than those of the stomach, and hence reflux of the stomach contents is prevented. Occasionally there is a drop in intra-esophageal pressure and reflux occurs. It's perceived symptomatically as "heartburn" in humans. If it persists the erosive effect of the acid stomach contents can cause ulceration of the lower esophagus; chronic irritation can lead to changes in the nature of the lining and eventually to cancer if untreated.
In most species the epithelial transition from esophagus to stomach is right about at the point where the anatomic esophagus and the anatomic stomach meet. But in others (such as the horse and rat) it occurs actually inside the anatomic stomach. Thus the gross anatomist will speak of the "non-glandular" and "glandular" regions of the stomach. In such situations the point of change is grossly visible, and the gross anatomist's term for it is the margo plicatus.
Gastric Mucosa: Gastric Pits and Mucosal Glands
Inside the actual stomach, the tunica mucosa differs in appearance and in function in different regions.
While in most of the digestive tract the lining epithelium is protective in
function (as, for example, in the esophagus) or absorptive (as in the
intestines), in the stomach the mucosal epithelium is secretory, and produces
products essential to the digestive process.
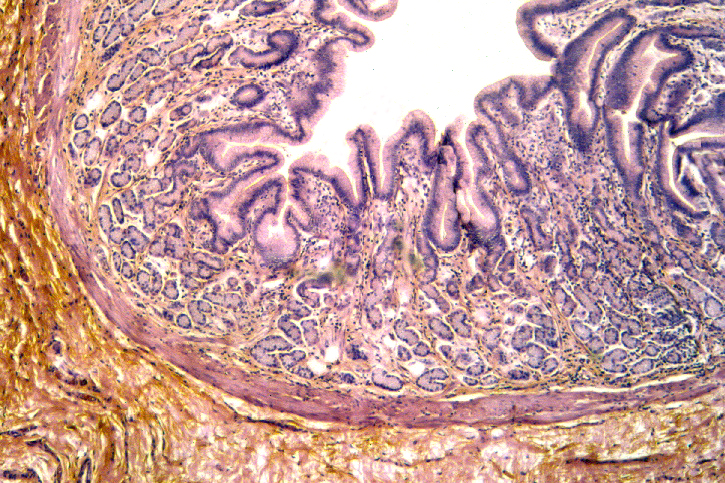 Using slide 133, you should be able to see that the lumen of the stomach is
lined with an ordinary looking simple columnar epithelium. There are no goblet
cells in it (in which respect it differs from the intestines). If you were
somehow able to stand inside the stomach and look at it en face, you'd
see deep depressions in the "floor" representing gastric pits or
foveolae, also lined with this simple columnar epithelium. Small
openings into the underlying regions of the mucosal tunic are found at the
bottom of these pits. If you took out your Swiss Army Knife and started
excavations into the lamina propria and the regions beneath the pits, you'd
find the actual secretory structures, the various types of gastric mucosal
glands.
Using slide 133, you should be able to see that the lumen of the stomach is
lined with an ordinary looking simple columnar epithelium. There are no goblet
cells in it (in which respect it differs from the intestines). If you were
somehow able to stand inside the stomach and look at it en face, you'd
see deep depressions in the "floor" representing gastric pits or
foveolae, also lined with this simple columnar epithelium. Small
openings into the underlying regions of the mucosal tunic are found at the
bottom of these pits. If you took out your Swiss Army Knife and started
excavations into the lamina propria and the regions beneath the pits, you'd
find the actual secretory structures, the various types of gastric mucosal
glands.
The image above shows this arrangement, and the scanning EM image at right gives a better impression of the open nature of the gastric lumen and the "hole-like" nature of the gastric pits. Note that in the stomach, in all its regions, the secretory glands are in the tunica mucosa, i.e., inside the muscularis mucosae. This is a key feature: the only two locations in which the glandular component of the digestive tract is outside the muscularis mucosae are the esophagus (see above) and the duodenum of the small intestine (see below). The glands open into the bottom of the pits.
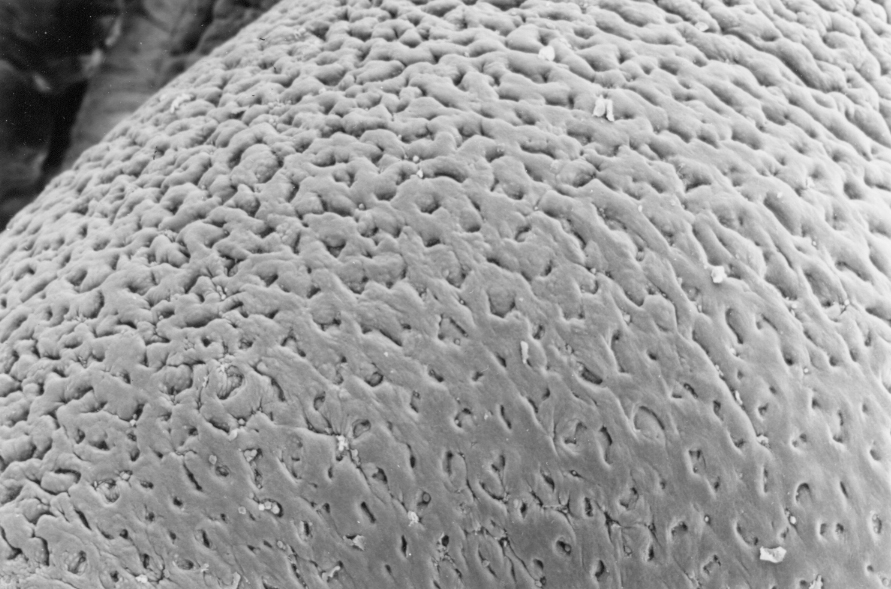 There are three major categories of these glands, associated with different
parts of the stomach. The glandular regions are wholly confined to the tunica
mucosa. Unlike some other portions of the tract, submucosal glands are not
found in the stomach at all.
There are three major categories of these glands, associated with different
parts of the stomach. The glandular regions are wholly confined to the tunica
mucosa. Unlike some other portions of the tract, submucosal glands are not
found in the stomach at all.
Gastric Glands of the Cardiac Region
The first category of gastric glands is the cardiac glands. These are found in the most
proximal region of the stomach (i.e., that part closest to the input of
the esophagus, called the "cardiac region," because it's located
close to the heart).
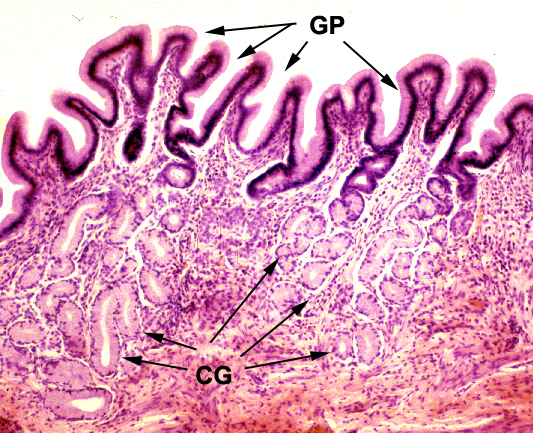 The gastric pits (GP) are lined with a simple columnar epithelium devoid of goblet cells. Cardiac glands (CG) make a mucous secretion for protection of the epithelium from erosion in the harsh environment of the stomach. At the bottom of the gastric pits are the openings into the secretory portions
of the glands. The glands themselves are tubular, extending deep into the
mucosa. They're limited in their
distribution to the stomach region immediately adjacent the esophagus, but occasionally
similar glands can be found in the esophagus itself. These are called "esophageal cardiac glands," and like the true cardiac glands of the stomach, they're in the tunica mucosa. This they're distinct from the esophageal glands proper, and probably represent individual
variations in development.
The gastric pits (GP) are lined with a simple columnar epithelium devoid of goblet cells. Cardiac glands (CG) make a mucous secretion for protection of the epithelium from erosion in the harsh environment of the stomach. At the bottom of the gastric pits are the openings into the secretory portions
of the glands. The glands themselves are tubular, extending deep into the
mucosa. They're limited in their
distribution to the stomach region immediately adjacent the esophagus, but occasionally
similar glands can be found in the esophagus itself. These are called "esophageal cardiac glands," and like the true cardiac glands of the stomach, they're in the tunica mucosa. This they're distinct from the esophageal glands proper, and probably represent individual
variations in development.
Gastric Glands of the Fundic Region
The second and by far the most numerous type of gastric gland is the fundic gland.
These are found underlying the bulk of the gastric mucosa. They produce
the bulk of secretions in the stomach and are best seen on slide 131. The
fundic glands, like the other types, open into the base of gastric pits. They
are deep, straight glands, with a mixed population of cell types.
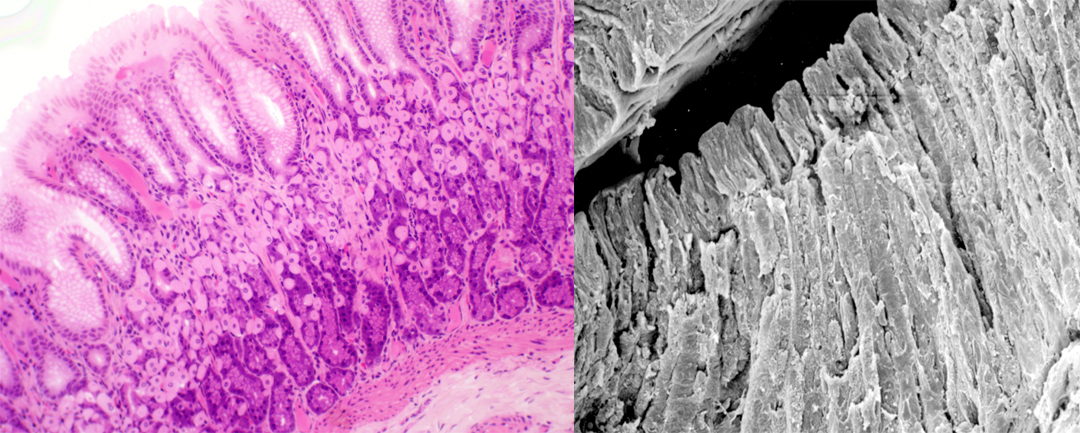
These two images are vertical sections through the fundic portion of the stomach, showing the relationship of the glands to the gastric pits above. On the left you see a low-power view through a number of glands. Even at this low magnification you can see there are several cell types in the deep region; we'll discuss them in detail below. At right the scanning EM image shows the deep pits opening to the surface and the compact regions of glandular tissue below, where the specimen was split apart. The fundic glands are by far the most numerous ones in mammalian stomachs. There are several million in most species. And as is true of all stomach glands, they're located in the deep parts of the tunica mucosa, surrounded by the connective tissue of the lamina propria.
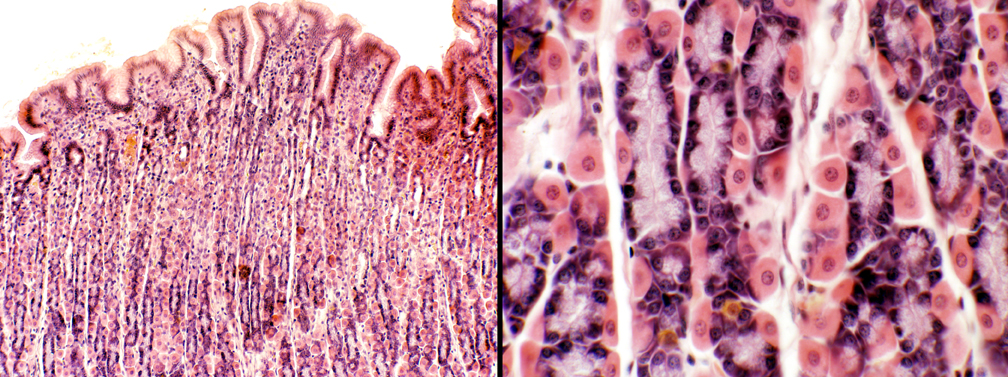
What isn't evident in either of these images is the structural category of the glands: they're simple branched tubular shape. The image on the right should this more plain.
This simple branched tubular gland assumes an inverted Y-shape: the "neck" of the gland is the stem of the Y and it is directed towards the top of the field (i.e., the free surface and the gastric pit).
At the point indicated by the arrows, the gland bifurcates into two long, tube-shaped secretory regions, in which reside the chief and parietal cells. These continue to the deepest levels of the lamina propria and butt up against the muscularis mucosae. They do NOT extend into the submucosal layer.
Cell Types in the Fundic Glands
The fundic glands are the source of the proteases of the stomach (trypsin and chymotrypsin) and the hydrochloric acid that gives the stomach contents its pH of about 2.0. In mammals these are produced by two different cell types: the parietal cells make the acid, and the chief cells make the proteases. In other groups of animals (e.g., birds) one cell type has both functions.
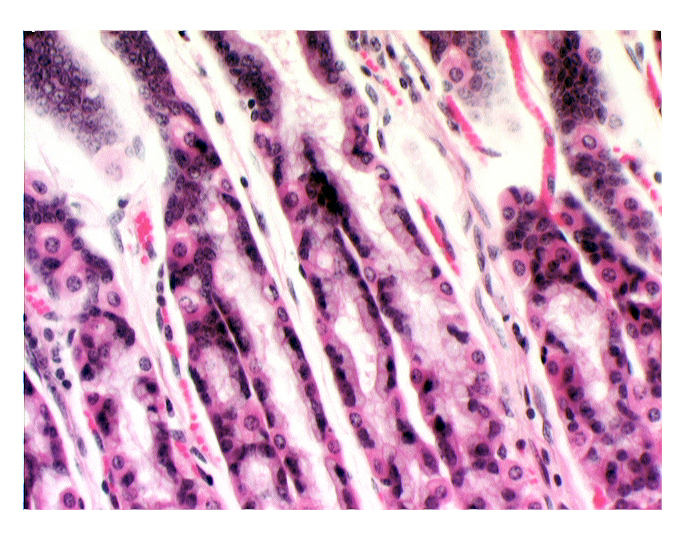
In this image the darkly stained cells are the chief cells. Chunky and angular in shape, and in H&E preparations very strongly basophilic, these cells have the typical 'secretory" appearance. In the electron microscope they show large amounts of RER and large numbers of secretory vesicles.
 The parietal cells are by contrast very eosinophilic in their staining response. and stand out against the background of the chief cells quite well. Their shape is different too: instead of being angular they tend to be rounded.
The parietal cells are by contrast very eosinophilic in their staining response. and stand out against the background of the chief cells quite well. Their shape is different too: instead of being angular they tend to be rounded.
The differences in staining and in shape are related to their functions. While the the chief cells show the typical properties of protein secreting cells, the parietal cells are essentially ion pumps. They have no protein secretion function and hence little RNA in their cytoplasm.
The parietal cells make hydrochloric
acid, to keep the pH of stomach juice low (about 2.0 to 3.0 is typical). This
pH is necessary to activate the 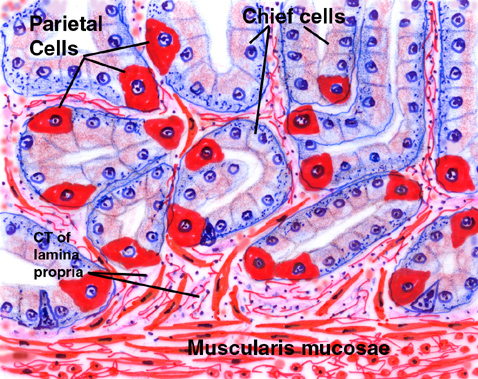 gastric enzymes. The role of the parietal cell
is to pump hydrogen ions out into the lumen of the stomach. It accomplishes
this task by extensive infolding of its apical surface and an enormous number
of mitochondria. The mitochondria inside the parietal cell can force hydrogen
ions across the membranes using active transport; this creates a sharp
concentration gradient between the surface channels and the open lumen of the
gland. The localized concentration gets so high that the ions diffuse out into
the lumen of the gland and eventually into the stomach.
gastric enzymes. The role of the parietal cell
is to pump hydrogen ions out into the lumen of the stomach. It accomplishes
this task by extensive infolding of its apical surface and an enormous number
of mitochondria. The mitochondria inside the parietal cell can force hydrogen
ions across the membranes using active transport; this creates a sharp
concentration gradient between the surface channels and the open lumen of the
gland. The localized concentration gets so high that the ions diffuse out into
the lumen of the gland and eventually into the stomach.
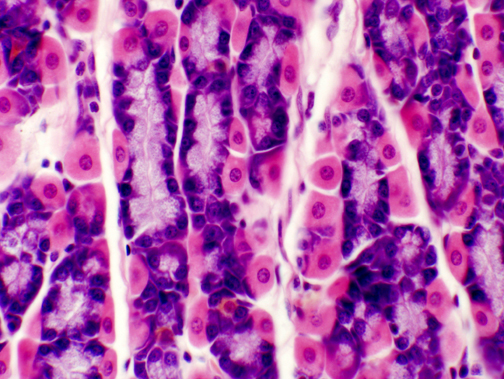
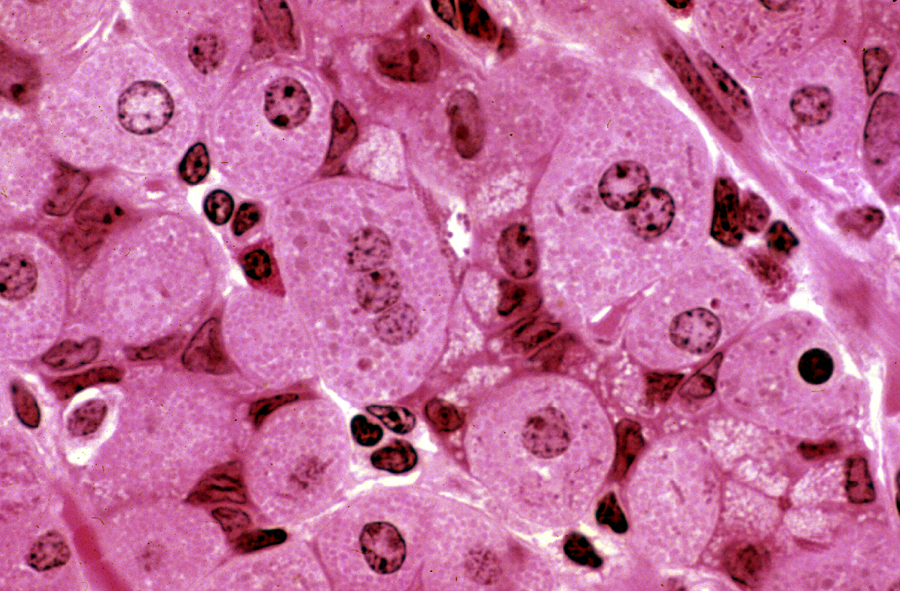 Parietal cells are often binucleated: and in this image you can see a tri-nucleated one. The role of the parietal cell is to pump hydrogen ions out into the lumen of the stomach, thus acidifying the contents. It is able to do so by virtue of having enormous numbers of mitochondria (to provide the energy needed for active ion transport), and a system of extensive apical invaginations into which the ions are passed, creating localized concentration gradients that favor diffusion into the lumen. In the image at left, the "granularity" of the cytoplasm reflects both these features.
Parietal cells are often binucleated: and in this image you can see a tri-nucleated one. The role of the parietal cell is to pump hydrogen ions out into the lumen of the stomach, thus acidifying the contents. It is able to do so by virtue of having enormous numbers of mitochondria (to provide the energy needed for active ion transport), and a system of extensive apical invaginations into which the ions are passed, creating localized concentration gradients that favor diffusion into the lumen. In the image at left, the "granularity" of the cytoplasm reflects both these features.
The chief cell is not nearly so dramatic in appearance, though it's equally important. Chief cells are shoved in among the parietal cells in groups of two or three. The enzymes produced by the chief cells are released in an inactive form; the acidic environment of the stomach hydrolyzes a fragment from the precursors, converting them to active enzymes.
Gastric Glands of the Pyloric Region
The third category of gastric glands is pyloric glands, found in the
region of the pylorus, the junction with the first part of the
intestine.
Like the cardiac glands, pyloric glands produce a mucous secretion. Structurally they closely resemble
the cardiac glands, though in the pyloric stomach the gastric
pits tend to be deeper and the glands larger and more obvious. Here you see the pyloric junction, the transition point between the stomach and the intestine.
 The stomach is a very hostile environment for cells; after all, it's a
place where very similar materials are in the process of being enzymatically
destroyed, and the pH is very low. To prevent too-rapid erosion of the gastric
mucosa, the surface cells and cells of the gland necks produce mucous
secretions which act to protect them. The protection isn't perfect, and there
is a constant shedding of dead cells from the gastric pits into the lumen. They
are replaced by new cells, moving up from the deep regions. As we'll see in
other regions of the digestive system, this is a common strategy: taking
advantage of the vast proliferative capabilities of epithelium to overcome
rapid erosion and damage. very large and open gastric pits are characteristic of this part of the stomach.
The stomach is a very hostile environment for cells; after all, it's a
place where very similar materials are in the process of being enzymatically
destroyed, and the pH is very low. To prevent too-rapid erosion of the gastric
mucosa, the surface cells and cells of the gland necks produce mucous
secretions which act to protect them. The protection isn't perfect, and there
is a constant shedding of dead cells from the gastric pits into the lumen. They
are replaced by new cells, moving up from the deep regions. As we'll see in
other regions of the digestive system, this is a common strategy: taking
advantage of the vast proliferative capabilities of epithelium to overcome
rapid erosion and damage. very large and open gastric pits are characteristic of this part of the stomach.
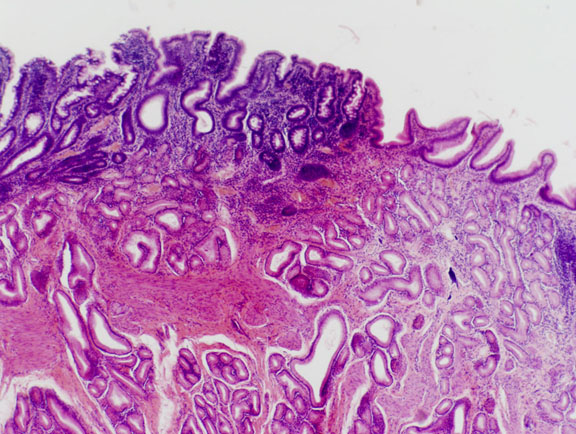
This image shows the pyloric region and the beginning of the duodenum. The mucosal glands of the pyloric region are similar in appearance to the glands of the cardiac region, and different from those of the fundic region. The overlying epithelium in the stomach is simple columnar with no goblet cells, whereas that of the duodenum has goblet cells, as does the rest of the intestine. Note also the appearance of gland profiles in the submucosa: this is another indication that we've reached the transition point.
Tunica Muscularis
The tunica muscularis of the stomach, unlike the bulk of the tract, is generally
considered to have three layers. While the layers are usually described as being oriented longitudinally, obliquely, and circumferentially with respect to the organ's long axis, this is really an oversimplification: they aren't very clearly defined and they intermingle with each other in places. The net effect of their contraction, however, is to cause the stomach to wring and twist itself about the long and short axes, so as to pulp up the ingested food and create the acidic chyme, consisting of the semi-liquified food and gastric secretions. That passes through the pylorus into the duodenum. Nervous elements in the submucosa and among the layers of the tunica muscularis coordinate and control the contractions of these muscles, as they do in other parts of the digestive tract.
 The most striking feature of the tunica muscularis externa of the stomach is the huge development of the pyloric sphincter, a muscular constriction that can open and close the pylorus to allow macerated food to pass. Unlike the "sphincter" of the esophageal-stomach junction, this is a true "valve" controlling flow.
The most striking feature of the tunica muscularis externa of the stomach is the huge development of the pyloric sphincter, a muscular constriction that can open and close the pylorus to allow macerated food to pass. Unlike the "sphincter" of the esophageal-stomach junction, this is a true "valve" controlling flow.
Some appreciation for the structure of this sphincter can be gained from the low-power image above, but the one at right really emphasizes it more. In this image the stomach region is to the right, the duodenum to the left, as in the images above. Only part of the tunica muscularis is shown, the large expansion of the middle layer that forms the bulk of the sphincter.
Myenteric Plexus
Muscles, even smooth muscles, need neural elements to control them. You will see
portions of the extensive neural network of the gut on almost any of the slides
examined thus far if you look between the muscle layers of the tunica
muscularis. This myenteric plexus (and a coordinated submucosal plexus) is present throughout the entire length of the digestive tract from the esophagus to the anus. (The submucosal plexus is much easier to see in the intestines, but it's present in the stomach as well.)
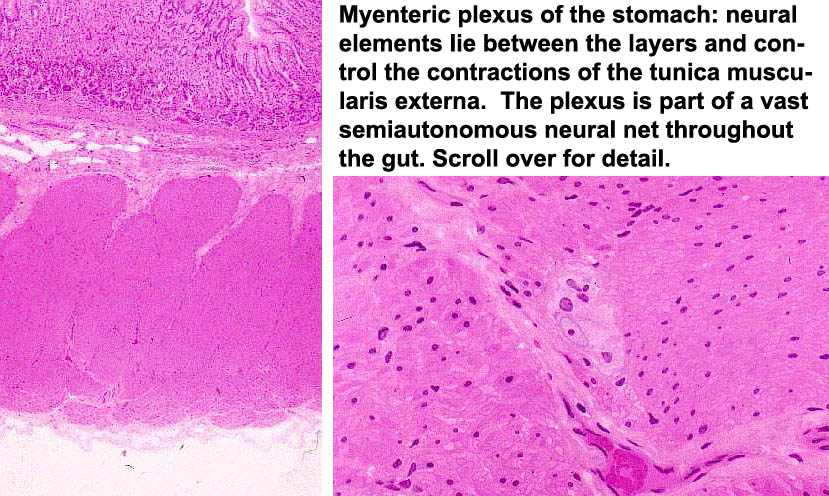
The myenteric plexus is a network of nerve fibers and neuron cell bodies, hence the plexus fulfills the definition of a "ganglion," a collection of neuron cell bodies outside the CNS. Note carefully that there is only one myenteric plexus that runs throughout the gut; the casual use of the plural in some texts implies that there are several, but it's really all one network. The smooth muscles of the tunica muscularis receive "orders" via these nerve fibers and contract in response. The plexus coordinates the contractions so that the appropriate kneading and squishing of the ingested food takes place. Here you see a knot of neurons, comprising one of the local nodes of the network.
The digestive tract thus has what amounts to its own division of the peripheral nervous system, sometimes called the "enteric nervous system," to distinguish it from the somatic and autonomic divisions. It is equivalent in complexity and autonomy to the others. Gastrointestinal sensory and motor activity pretty much runs on "autopilot" except when there is some overwhelming need for the CNS to get involved.
Lab Exercise List