Objectives for This Exercise
Objectives for This ExerciseSLIDES FOR THIS EXERCISE: 40, 41, VM77, 849, VM79, VM82, VM80, VM81
PRE-RECORDED LECTURE FOR THIS EXERCISE
The intestines are the parts of the digestive system responsible for the absorption of nutrients and water. Two anatomic regions exist, the small intestine and the large intestine. Both of these are further subdivided into anatomically discernible subdivisions. The small intestine has three parts: the duodenum, the jejunum, and the ileum; the large intestine is subdivided into the colon, cecum, rectum, and is continuous with the anus, the last portion of the alimentary canal.

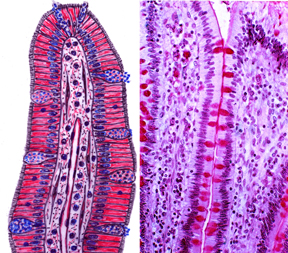
This diagram lays out in schematic form some of the salient features of the two major subdivisions of the intestine, and highlights their similarities and differences. These are key things that will enable you to distinguish them from each other in the microscope. Overall the intestines have the normal form expected of tubular organs, including the four "tunics" discussed in Exercise 18, the tunica mucosa, tunica submucosa, tunica muscularis externa, and the tunica serosa.
 The most visible and significant difference is the presence of villi in the small intestine, and their absence in the large intestine. The small intestine is a place in which nutrients are absorbed, and it has a huge surface area to fulfill this role (see below). Material moving through here is in liquid form. The villi are a means to enhance the absorptive surface, and the contents can flow around and over them efficiently. between the protruding villi are deep intestinal crypts where new epithelial cells are generated by mitosis.
The most visible and significant difference is the presence of villi in the small intestine, and their absence in the large intestine. The small intestine is a place in which nutrients are absorbed, and it has a huge surface area to fulfill this role (see below). Material moving through here is in liquid form. The villi are a means to enhance the absorptive surface, and the contents can flow around and over them efficiently. between the protruding villi are deep intestinal crypts where new epithelial cells are generated by mitosis.
The large intestine primarily absorbs water, and compacts and dries out the fecal bolus: villi would be a hindrance to movement of the semi-solid fecal mass, and would likely be injured by its passage. hence there are no villi in the large intestine, and in addition, there are numerous goblet cells whose secretions act as lubrication for the moving material.
Many people have the erroneous idea that the intestines actually secrete digestive enzymes (you'll actually hear this implied on some TV commercials for diet products and supplements). They don't. It's the pancreas which produces the digestive enzymes used for the process; we'll look at that in another exercise.
Actual digestion or breakdown of foodstuffs in the gut is extracellular; i.e., it takes place in the lumen of the tube, with the resulting breakdown products being absorbed across the plasma membrane of the lining cells. This process represents a nice application of a general rule: if you want to improve the efficiency of a process which is associated with or limited by a transmembrane phenomenon, the way to do it is increase the surface area.
Consider a hypothetical small intestine with a length of 5 meters and a diameter of 25 mm. This tube, without modification, would have a volume of a little less than a liter, and a surface area of 3.9x106 square millimeters, approximately the size of a small windowpane. Increasing the total surface area available produces an enormous increase in the efficiency of the absorptive process, since the rate of absorption per unit area is limited.
If an average villus is 1.0 mm long by 0.5 mm in diameter, and if villi are distributed at a uniform 25 per square millimeter, the increase in surface area compared to a simple tube is substantial. With villi present at this density, the available surface area for absorption from that volume would be more on the order of a very large room. Since the villi are not the only (or even the principal) adaptation to increase the surface area, the actual increase is vastly greater: the presence of microvilli on the cells and crypts between villi increases the surface area available for absorption to several acres. This enormous reserve capacity is essential. Loss of some of it to injury, scarring, surgical excision, etc., leaves plenty of functional tissue to allow for normal nutritional intake.
Need more absorption capability? Then make more absorptive surface. This design solution is found over and over in the "engineering" of living organisms.
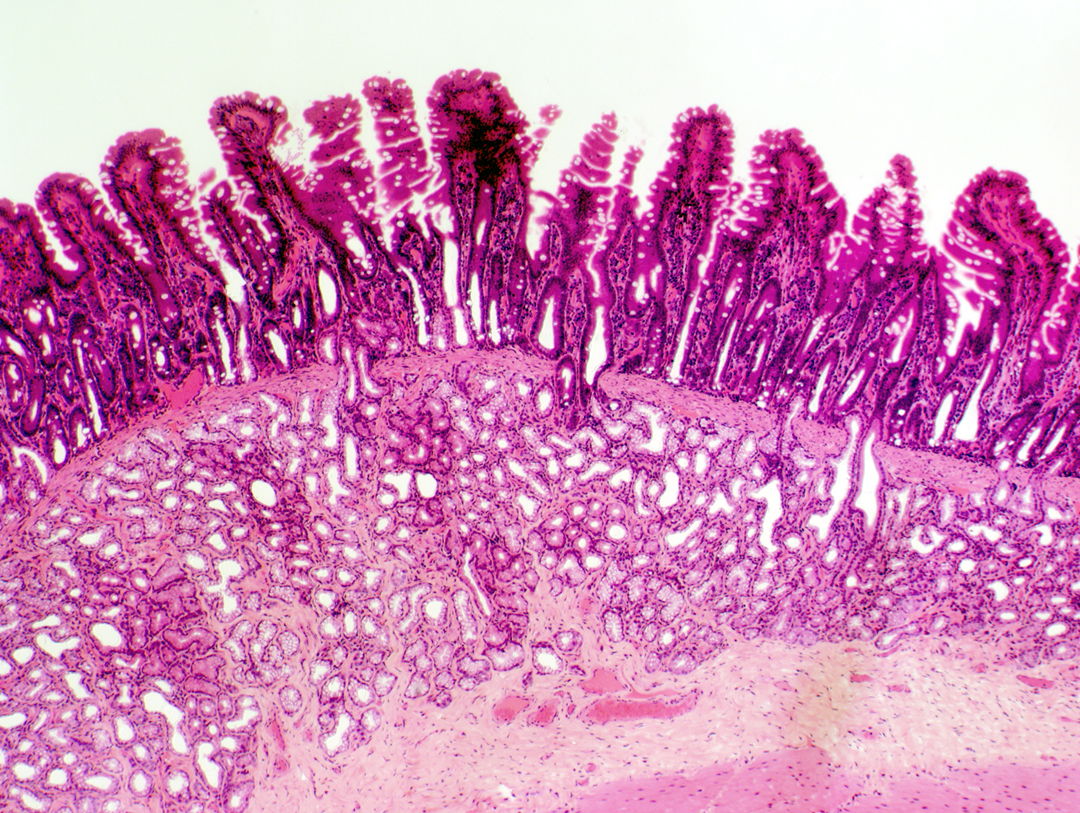
Begin with slide 40, a piece of duodenum (from the Latin for "twelve," so named in medieval times because in humans it's twelve finger breadths in length), the first portion of the small intestine. Most of the general structure of the intestine is similar throughout, and the overall description given here for duodenum is appropriate for the jejunum and the ileum, with some specific regional variations that will be described below for all three regions. These structural variations serve as "landmarks" in the histological identification of different areas.
The lamina propria , the delicate CT that fills the cores of the villi and the spaces between the crypts, is demarcated from the more peripheral layers by the muscularis mucosae, a thin band of smooth muscle which runs all the way around the mucosal layer and which sends strands up into the villi. It's believed these strands cause the villi to contract, expelling the contents of the crypts and the intervillous spaces. The muscularis mucosae is the outermost and last layer of the tunica mucosa of the intestine. The low power view above shows the general appearance of the duodenum. Only part of the tunica muscularis externa is visible in this field.
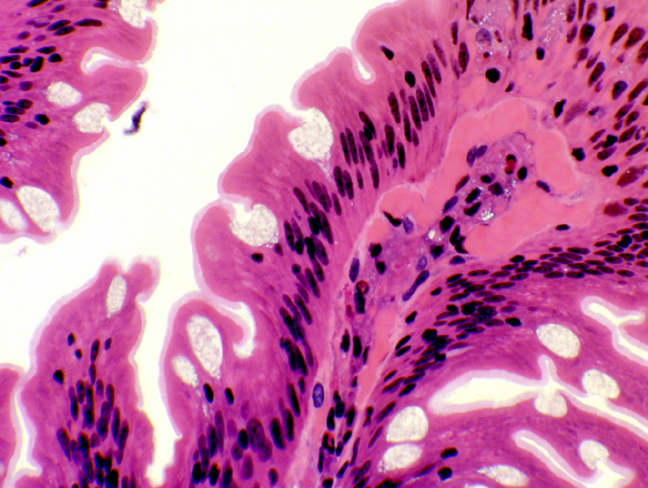 Notice there are several places in here where the glands' ducts are breaking through the muscularis mucosae to dump their secretions into the deepest regions of the intestinal crypts. Unlike the stomach, the crypts (which are called "glands" in some
texts) do NOT secrete digestive enzymes or hydrochloric acid.
Notice there are several places in here where the glands' ducts are breaking through the muscularis mucosae to dump their secretions into the deepest regions of the intestinal crypts. Unlike the stomach, the crypts (which are called "glands" in some
texts) do NOT secrete digestive enzymes or hydrochloric acid. 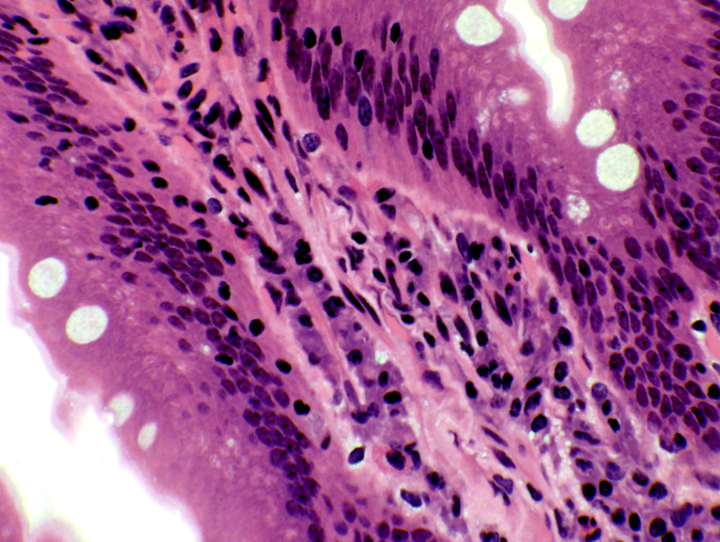 The duodenal mucosa (and that of the rest of the intestines) is a sheet of simple columnar epithelium. The cells rest on a basal lamina over the lamina propria, the loose collagenous CT, filled with cells, that constitutes the core of each villus.
The duodenal mucosa (and that of the rest of the intestines) is a sheet of simple columnar epithelium. The cells rest on a basal lamina over the lamina propria, the loose collagenous CT, filled with cells, that constitutes the core of each villus. 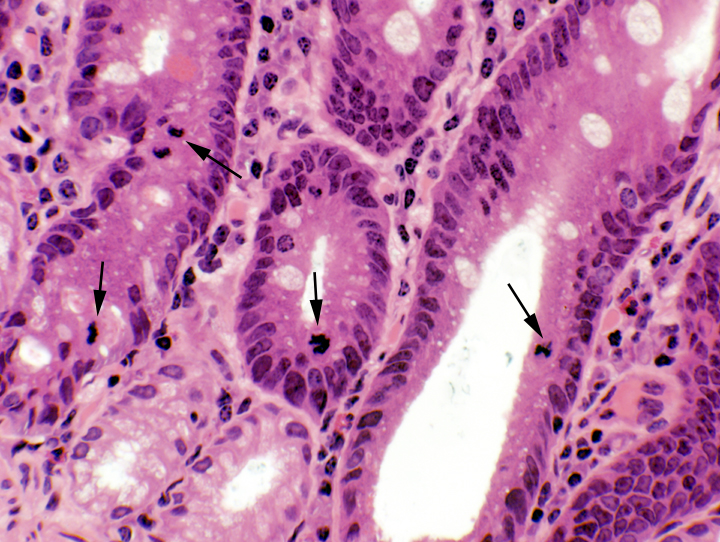 Constant proliferation of enterocytes and goblet cells in the deepest portion of the crypts replaces the vast numbers shed at the apex of each villus. Examining the deep regions of the crypts will show many mitotic figures (right).
Constant proliferation of enterocytes and goblet cells in the deepest portion of the crypts replaces the vast numbers shed at the apex of each villus. Examining the deep regions of the crypts will show many mitotic figures (right).Cell Types in the Tunica Mucosa
Most of the cells in the digestive tract are the absorptive enterocytes, but there are other types present.
Goblet Cells
 Throughout the intestinal tract from the duodenum distally, numerous secrete a mucoid material which slows down the loss of the enterocytes. The PAS stain and any other stain for carbohydrates will make goblet cells stand out against the far more numerous enterocytes. Goblet cells aren't found in the stomach. While they're very numerous in the small intestine, they are even more so in the large intestine, thanks to the lubricating properties of their mucus (see below)
Throughout the intestinal tract from the duodenum distally, numerous secrete a mucoid material which slows down the loss of the enterocytes. The PAS stain and any other stain for carbohydrates will make goblet cells stand out against the far more numerous enterocytes. Goblet cells aren't found in the stomach. While they're very numerous in the small intestine, they are even more so in the large intestine, thanks to the lubricating properties of their mucus (see below)
Enteroendocrine Cells
Less numerous than goblet cells, but still very important, are the enteroendocrine cells. These can be seen on slide 40 as well. They're similar to those in the stomach, but produce different hormones. The entire gastrointestinal tract is in fact a large endocrine organ, with a diffuse and variegated population of these cells whose hormones signal a number of activities related to digestive physiology. At last count at least 20, perhaps as many as 40, such cells exist, each making a different hormone. They can't really be told apart morphologically with simple stains, but immunolabeling for their products can sort them out.
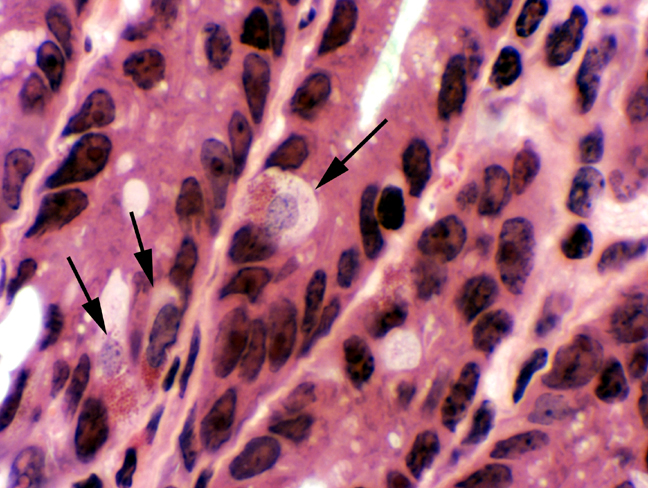 Look in the deep regions of the crypts for cells with granules in their cytoplasm, in which the granules are polarized
away from the lumen of the crypt and towards the lamina
propria. These cells don't secrete into the lumen; like any
self respecting hormone producing cell, they discharge their
secretions into the blood flowing through capillaries in the lamina
propria.
Look in the deep regions of the crypts for cells with granules in their cytoplasm, in which the granules are polarized
away from the lumen of the crypt and towards the lamina
propria. These cells don't secrete into the lumen; like any
self respecting hormone producing cell, they discharge their
secretions into the blood flowing through capillaries in the lamina
propria.
The functions of many of these endocrine cells have been established. Actually, their presence has long been known to histologists, but only in the past couple of decades has it been possible to establish their variety and their individual functions. In the image above an enteroendocrine cell has been revealed by a classic silver deposition method. Because these cells stain strongly with this sort of method, they have long been called "argentaffin cells" ("silver affinity"). The reaction precipitates metallic silver as 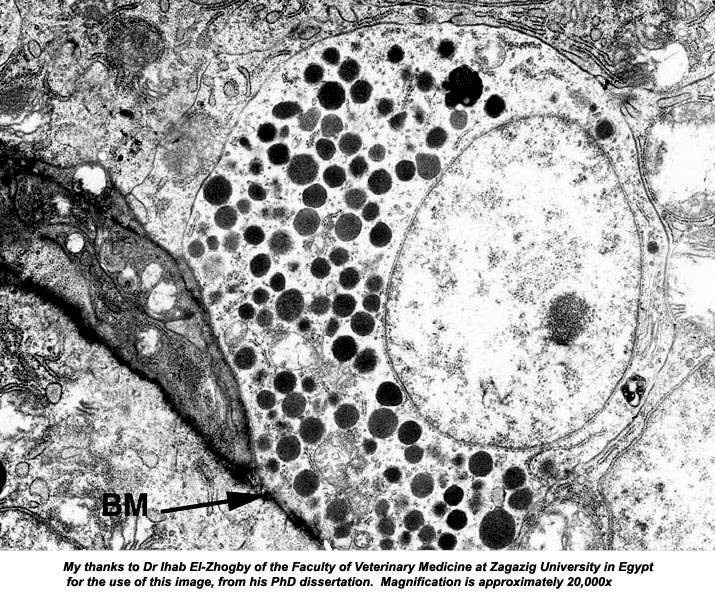 dark granules.
dark granules.
This electron microscope image demonstrates the nature and polarization of an enteroendocrine cell nicely. The round cell is located right on the basement membrane (BM) and its granules are located on the side below the nucleus and against the membrane. The nucleus of this cell is vesicular, and there's a prominent nucleolus. That plus the numerous secretory vesicles in the cytoplasm are evidence of its function.
There's no way to tell from an image like this one (or the light micrograph above) what the cell is producing; but immunolabeling studies have shown that enteroendocrine cells have target organs in distant regions of the digestive system. One is known to stimulate secretion in the pancreas, another to cause contraction of the smooth muscle in the wall of the gall bladder; and others target specific cells of the endocrine system itself. There's considerable evidence that many enteroendocrine cells can discharge in response to nervous stimulation, and that in some instances nerve fibers innervate them directly. Thus we have one more example of how the two systems for response to stimulus are integrated.
Paneth Cells
While you're hunting for enteroendocrine cells you'll undoubtedly
see another cell type on slide 40. In the depths of the crypts,
you will find clusters of 3 to 5 cells with coarse red
staining granules at the apical end. Note that these
cells are polarized towards the lumen. They are Paneth
cells, named for 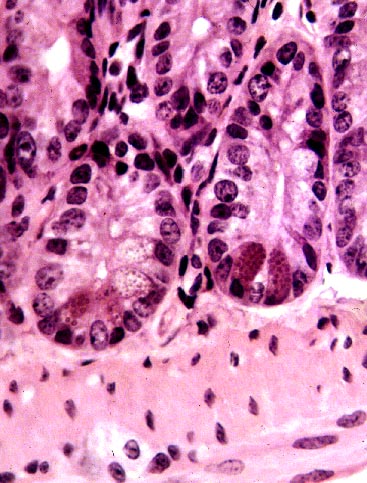 Josef Paneth (1857-1890), an Austrian physician
who discovered them more than a century ago. They are believed
to produce bactericidal materials and release them into the lumen of
the gut. Note that the Paneth cells aren't part of the
enteroendocrine system because they don't release hormones into the
blood, they release it into the gut lumen. Note the difference
in color between the Paneth cell granules and the granules in the
enteroendocrine cells. Paneth cells are found in the small intestine but not in the large intestine.
Josef Paneth (1857-1890), an Austrian physician
who discovered them more than a century ago. They are believed
to produce bactericidal materials and release them into the lumen of
the gut. Note that the Paneth cells aren't part of the
enteroendocrine system because they don't release hormones into the
blood, they release it into the gut lumen. Note the difference
in color between the Paneth cell granules and the granules in the
enteroendocrine cells. Paneth cells are found in the small intestine but not in the large intestine.
For many years the Paneth cell was a mystery whose function(s) were unknown for a century after it was first described. They're much more numerous in some species than others, and they're exceptionally abundant in some South American anteaters—leading to a speculation that they might have chitinase activity to digest the exoskeleton of arthropods, and attractive hypothesis that didn't stand up under testing. It's recently been shown that the granules contain a form of bactericide, and lysozyme-like agents. This suggests they're responsible for protecting the gut against bacterial overgrowth, and this is the current explanation of what they do—subject, of course, to revision as new data is gathered.
The granules of Paneth cells are much coarser and more obvious than those of enteroendocrine cells. The cells are usually found clustered in groups of three to six or so, located at the very deepest part of an intestinal crypt. Unlike the enterocytes and goblet cells, they're a very stable and long-lived population, and they don't migrate from the deep part of the crypts to the surface. They appear not to engage in mitosis. Some animals lack them, including dogs, pigs, cats, and raccoons.
The tunica submucosa is the region of CT immediately outside the muscularis mucosae. It has fair numbers of blood vessels and lymphatics in it, too, and if you look carefully you will be able to distinguish localized collections of neuron cell bodies. These are elements of the submucosal plexus discovered by Georg Meissner (1829-1905), a German histologist. This plexus, together with another one located in the tunica muscularis (see below) helps to coordinate the movements of the intestine and facilitate the passage of food through its lumen.
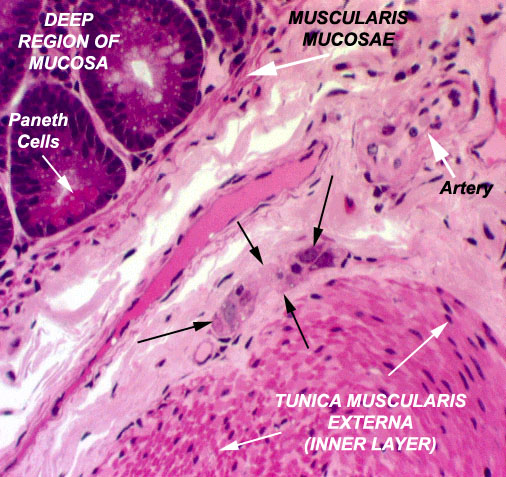
One of the collections of neurons of the submucosal plexus is shown in the image at left. The plexus is tucked into the interstices of the CT that is outside the muscularis mucosae.
It contains autonomic motor neurons and their attendant satellite cells, nerve fibers that connect the neurons to each other, and carry motor commands to effector cells. There are also fibroblasts that hold the structure together by forming a capsule and stromal elements. In this image a small artery is also labeled; and between the plexus and the deep region of the crypts there's a small vein cut longitudinally (unlabeled). By definition, this is a ganglion; its nodal points control the muscles of the muscularis mucosae (MM) and the strands of smooth muscle that run through the cores of the villi. Pulsatile contraction of the innermost part of the mucosa, and the movements of the villi, contribute to mixing and overturn of the food, and emptying of the mucosal crypts.
Nervous signals running through the submucosal plexus are coordinated with the signals sent out from the larger myenteric plexus (see below) which controls the larger movements of the gut wall, and peristalsis. The submucosal and myenteric plexuses together constitute an autonomous "enteric division" of the nervous system with its own sensory and motor arcs and feedback loops.
Myenteric Nerve Plexus
The tunica muscularis is quite thick, and in the duodenum (as well as in the rest of the intestines) has the typical two layers, inner circular and outer longitudinal.
Between these two layers is the second nervous plexus, of the intestine, the myenteric plexus (formerly called the "plexus of Auerbach," after Leopold Auerbach (1828-1897). You will see this plexus as discrete collections of neuron cell bodies, located at fairly regular intervals between the inner and outer muscle coats. If you could peel away the outer longitudinal muscle coat you'd see the myenteric plexus as a net-like structure, with axonal fibers extending from one "knot" of neurons to another.
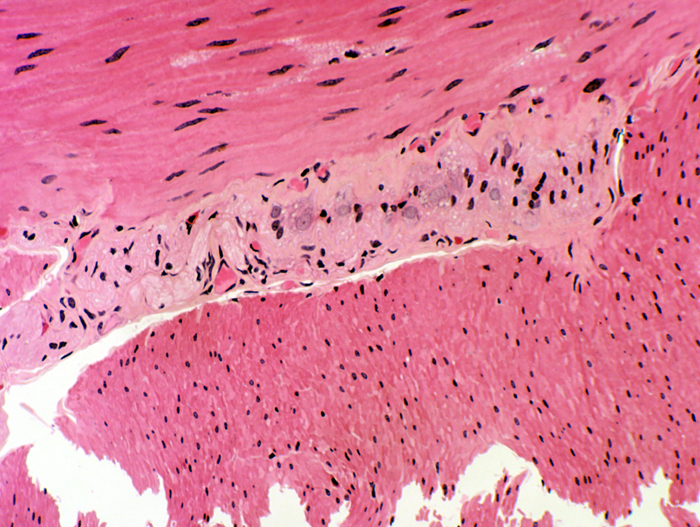 This image shows the relationship of the plexus and the tunica muscularis externa of the small intestine as it appears in the light microscope: there is an inner circular layer of smooth muscle (whose fibers are wrapped around the long axis of the tube); the outer layer that covers it has fibers running longitudinally.
This image shows the relationship of the plexus and the tunica muscularis externa of the small intestine as it appears in the light microscope: there is an inner circular layer of smooth muscle (whose fibers are wrapped around the long axis of the tube); the outer layer that covers it has fibers running longitudinally.
The space between them contains the myenteric plexus, whose function is to coordinate their peristaltic contraction.
Peristalsis requires that the wall of the tube be locally constricted and shortened; the circular and longitudinal layers are coordinated to do this. The constriction of the tube followed by its longitudinal shortening pushes the food bolus along in the proper direction.
In the small intestine the outer longitudinal layer is continuous over the surface, but in the large intestine, this isn't the case. The longitudinal bundles are still present but they're gathered into flat, narrow ribbons of muscle that lie spaced around the circumference of the tube. These are called the taenia coli, the "colonic ribbons," by the gross anatomist. There may be two or three of these, depending on the species.
The "Enteric Division" of the Autonomic Nervous System
If you trace the connections of the myenteric plexus (there are techniques to do this) you'd find that it's "wired" to the submucosal plexus. Each keeps the other "in the loop," about its activity, forming what is really a single functionally integrated system of neuronal elements, embedded in the intestinal wall at two levels. Operating together they comprise what is in effect an independent subdivision of the autonomic nervous system, the so called "enteric division."
The enteric nervous network consists of complete feedback loops and is pretty much capable of operating on its own without much "interference" (perhaps "guidance" is a better word) from the CNS. If a piece of living intestine is surgically removed and held in an appropriate bath, stimulation of the villi by physical touch causes them to shorten, and distention of the tunica muscularis causes contraction in response, entirely without any "oversight" by the CNS. In vivo the tract detects the presence of food, sends signals to the muscle fibers of the tunica muscularis and the muscularis mucosae, and under normal conditions processes the food through the duodenum, jejunum, ileum, and the large intestine more or less automatically.
The submucosal plexus is believed to control the fibers of the muscularis mucosae and the villi, to cause shortening of the villi and a "squishing out" of nutrients in the crypts and between villi. The myenteric plexus is responsible for the coordination of the rhythmic contract-shorten-contract-shorten movement of the tunica muscularis, that produces peristalsis. The two work in communication and coordination with each other.
Of course, as is discussed in Exercise 9, there is really only one "nervous system," however many functional divisions it may have, and the brain is always in command of all of it. If necessity demands it, the peristalsis can be stopped by CNS input or hormonal action triggered by CNS activity.
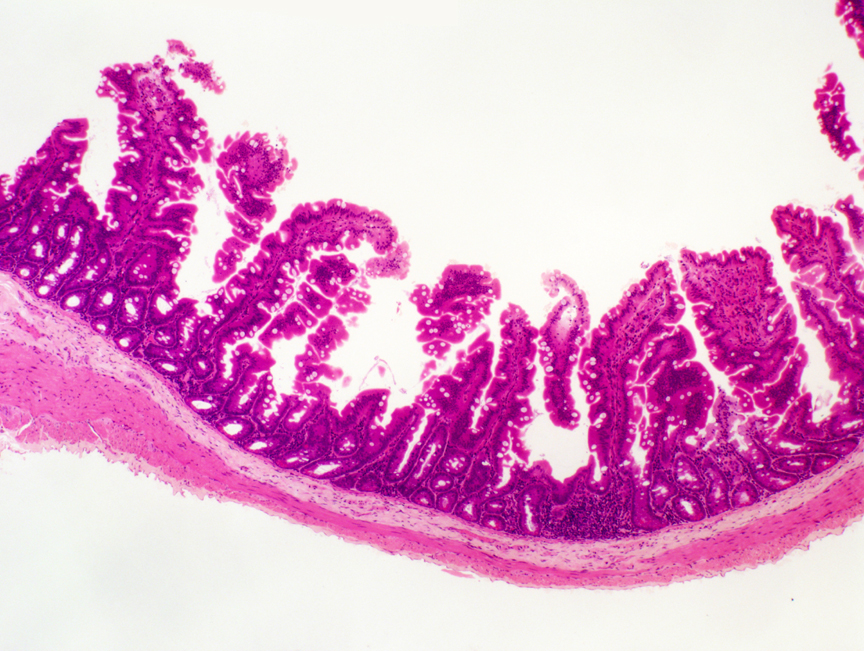 The next region of the small intestine is the jejunum(from the Latin word for "empty"), which you'll see on slide 124. Jejunal villi are longer and more irregular than the short, blunt
villi characteristic of the duodenum. It's in this section that
the bulk of absorption of nutrients takes place, and so you'd expect
the ramification of surface area to be extensive, and it certainly
is.
The next region of the small intestine is the jejunum(from the Latin word for "empty"), which you'll see on slide 124. Jejunal villi are longer and more irregular than the short, blunt
villi characteristic of the duodenum. It's in this section that
the bulk of absorption of nutrients takes place, and so you'd expect
the ramification of surface area to be extensive, and it certainly
is.
Notice that in this example there are what appear to be "detached" villi here and there, "floating" between those anchored to the bottom of the section. Villi these are, detached they are not.
These "islands" are often seen in sections of jejunum, but not in the duodenum: villi in the jejunum are very long, several millimeters, much longer than those of the duodenum. The section rarely will exceed 7 microns in thickness. When you're slicing up a round, tube-like structure with villi projecting into it from all sides, the tops of the villi that are longer than the section is thick will project into the section plan. A serial section through this area would prove beyond doubt that all the "islands" are attached outside the section plane, and that the "floating" appearance is spurious.
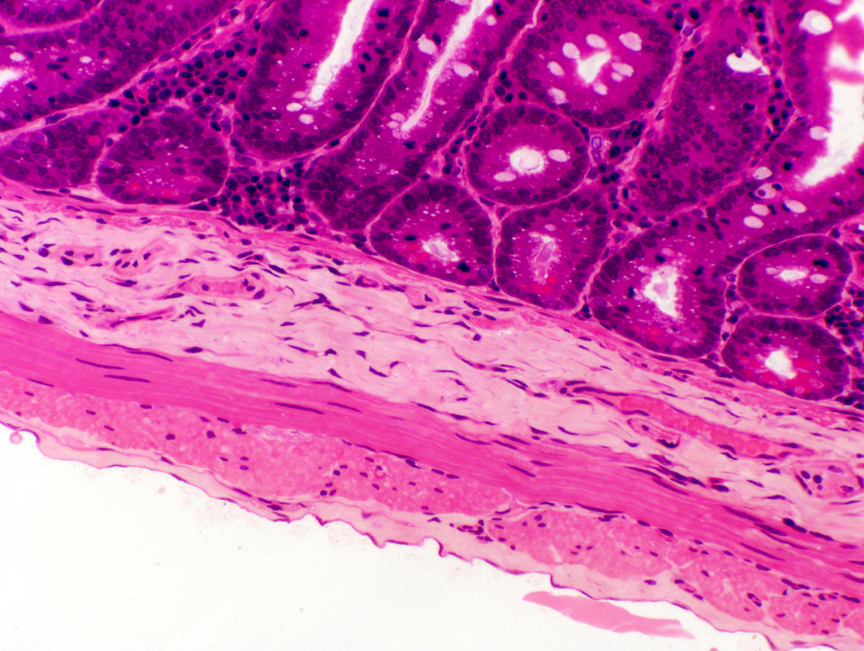 These "islands" are a visual clue that this isn't a section of duodenum. Another clue is the absence of structures in the submucosa. If this were duodenum, you'd expect to see submucosal glands; if it were ileum, aggregated lymphatic nodules should be present. Neither is...so, by default, this is jejunum.
These "islands" are a visual clue that this isn't a section of duodenum. Another clue is the absence of structures in the submucosa. If this were duodenum, you'd expect to see submucosal glands; if it were ileum, aggregated lymphatic nodules should be present. Neither is...so, by default, this is jejunum.
The muscularis mucosae here is sparse or even absent. Gross folds which involve not only the tunica mucosa., but also the underlying submucosa, are common. These folds, the plicae circulares, are permanent structures, another means for increasing surface area. Other features are the same as those in the duodenum. Notice that there are no glands in the submucosa in this region.
The ileum is the next portion (from the Greek, "eileo," to
roll up or twist). It's on slide 841. Most of the features you will see in this portion of the intestine
are similar to those in the previous sections, but the villi are even
more leaflike and you probably will see "islands" apparently floating
free in the 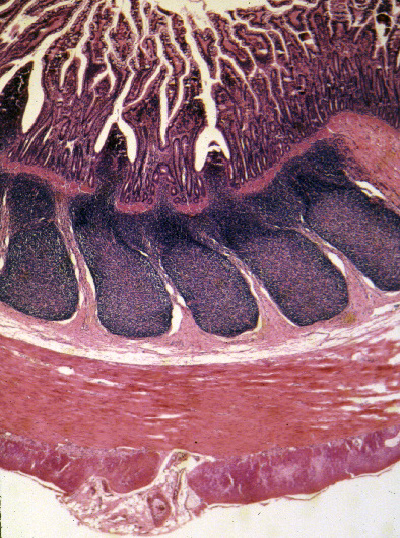 lumen. The most prominent landmark in the ileum is the presence of large
aggregations of lymphatic tissue in the submucosa. These are
the aggregated lymphatic nodules (usually called Peyer's
Patches after Johannes K. Peyer (1653-1712), a Swiss anatomist).
These can become so large under some situations that they'll infiltrate past the
muscularis mucosae and up into the mucosa proper, sometimes
obliterating the enterocytes.
lumen. The most prominent landmark in the ileum is the presence of large
aggregations of lymphatic tissue in the submucosa. These are
the aggregated lymphatic nodules (usually called Peyer's
Patches after Johannes K. Peyer (1653-1712), a Swiss anatomist).
These can become so large under some situations that they'll infiltrate past the
muscularis mucosae and up into the mucosa proper, sometimes
obliterating the enterocytes.
The submucosal lymphatic nodules frequently contain germinal centers. They represent the site of maturation and development of the B-lymphocytes involved in humoral immunity. While they're not transient structures, they do vary in number depending on the age of the animal. This image shows its characteristic features clearly, especially the aggregated lymphatic nodules. The nodules are the mammalian equivalent of the bursa of Fabricius in birds. They're most prominent in growing animals who are expanding their repertoire of immune responses, and tend to get smaller in aged ones. However, they're never lost completely. Note that Peyer's Patches are in the submucosa. That is, they are outside the muscularis mucosae, between it and the tunica muscularis. The tunica muscularis of the ileum has the typical inner circular and outer longitudinal layers.
Diffuse lymphatic tissue and localized concentrations of lymphocytes can occur almost anywhere in the gut. Such transient and structurally unorganized collections aren't the same thing as what's shown here. Peyer's Patches are organized, well-defined structures, a constant feature of the ileum in all mammals and most other vertebrates. They're "nodular" lymphatic tissue, and often will contain germinal centers. Germinal centers never occur in randomly-forming, transient aggregations.
Since the function of the large intestine is to desiccate and compact and lubricate the fecal bolus, while the layers present are similar to those of the small intestine, villi are absent; there are only crypts. The number of goblet cells is greatly increased compared to the small intestine as well. The tunica muscularis is scantier, and there may well be some small lymphatic nodules present.
Colon & Rectum
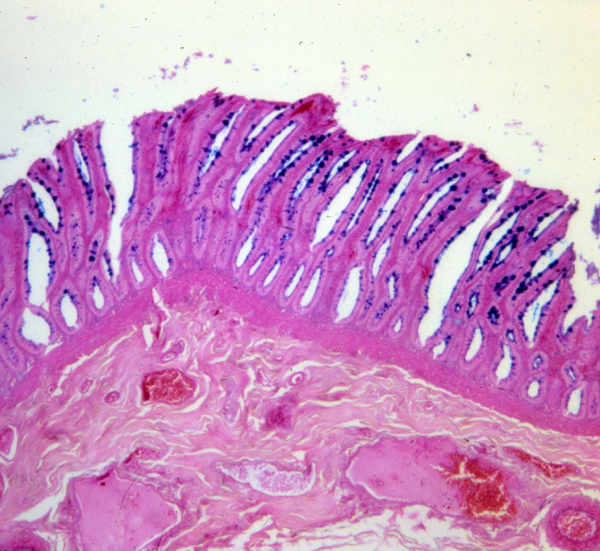 The colon is seen on slide 126. The large intestine is characterized by deep, straight crypts in which are found large numbers of goblet cells. Since most nutrients have already been extracted, surface area enhancement isn't as important as it is in the small intestine, but when you're moving large amounts of relatively dry material,what you need is lubrication, and a good deal of it.
The colon is seen on slide 126. The large intestine is characterized by deep, straight crypts in which are found large numbers of goblet cells. Since most nutrients have already been extracted, surface area enhancement isn't as important as it is in the small intestine, but when you're moving large amounts of relatively dry material,what you need is lubrication, and a good deal of it.
Obviously the drier the bolus gets, the harder it is to shove along. The large numbers of goblet cells ease the efforts of the tunica muscularis. The crypts are much more open to the lumen than those of the small intestine, which is to be expected. The lamina propria underlying the lumen fills the spaces between the crypts.
The crypts of the large intestine are filled with vast numbers of goblet cells that secrete slimy mucus to lubricate the poop to facilitate its...um...ejection. This is clear from the PAS-stained image shown here. Easy access to the lumen is needed for the cell product to reach the surface to coat the moving material.
Now...imagine what would happen if: 1) water weren't resorbed efficiently in the large intestine; or, conversely, 2) water were resorbed and goblet cell activity diminished or stopped. Neither situation is a happy one to contemplate, but sooner or later every dog owner has to deal with both.
The colon is one of those places where neoplastic lesions tend to develop. Colon cancer is bad news, indeed: but with periodic monitoring it is completely preventable.
The rectum is the last part of the colon, and it's distinguished by the presence of an enormously enlarged tunica muscularis, forming the anal sphincter.
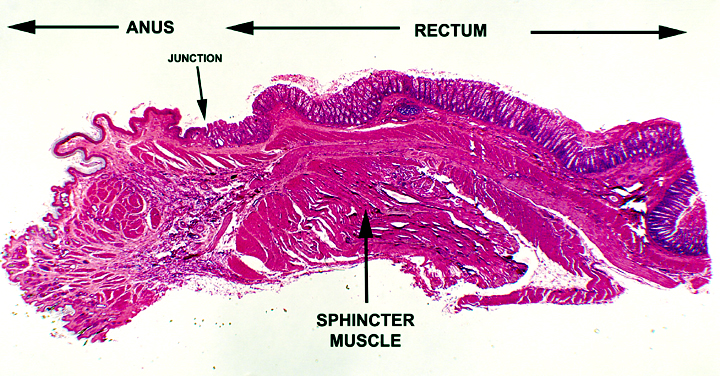
The anorectal junction, where these two portions of the
tract come together, is seen on slide 1209. While the rectum
forms from embryonic endoderm, the anus, is derived from
surface ectoderm. 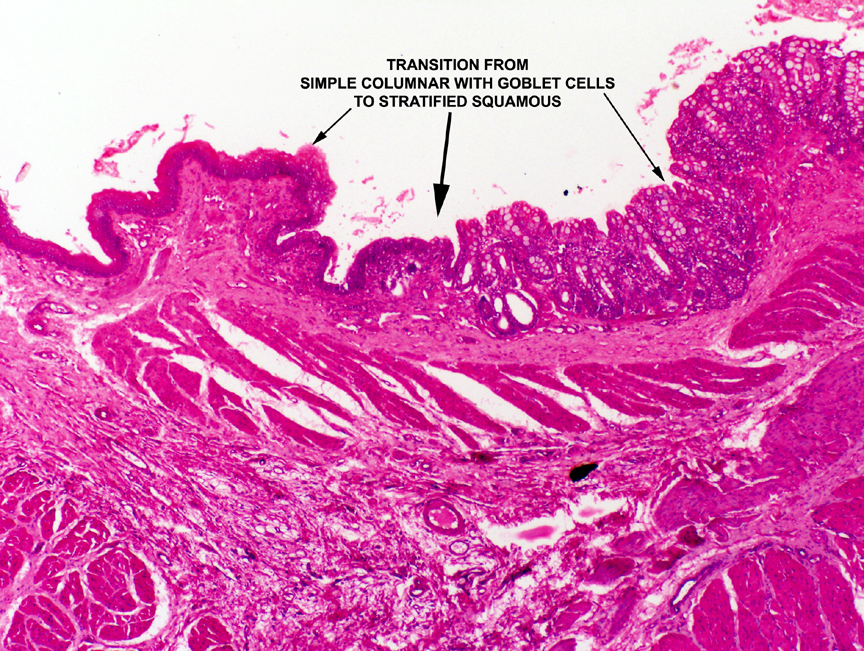 Therefore, the anus, the last few centimeters
of the digestive tract, is lined not with simple columnar epithelium,
but with stratified squamous, usually keratinized. The transition
point is easily made out on this slide, and note how very abrupt it
is. Just as it is at the oral end, the changeover from one type
of epithelium to another is very noticeable. This region of the tract has an abrupt transition of epithelial type from simple columnar in the rectum to stratified squamous in the anal part. The point of change over is marked in the image on the right. Of course, in addition to the change of epithelial type, there's an immediate disappearance of the crypts and goblet cells.
Therefore, the anus, the last few centimeters
of the digestive tract, is lined not with simple columnar epithelium,
but with stratified squamous, usually keratinized. The transition
point is easily made out on this slide, and note how very abrupt it
is. Just as it is at the oral end, the changeover from one type
of epithelium to another is very noticeable. This region of the tract has an abrupt transition of epithelial type from simple columnar in the rectum to stratified squamous in the anal part. The point of change over is marked in the image on the right. Of course, in addition to the change of epithelial type, there's an immediate disappearance of the crypts and goblet cells.
Remember that there is a similar site at the cardiac junction, where the esophageal epithelium (stratified squamous) abruptly transits to the epithelium of the stomach (simple columnar). How do you tell these two junctions apart? Look at the columnar epithelium. If it has goblet cells in the sheet, it's the ano-rectal junction. Another giveaway is the deep straight crypts of the rectum, and the absence of glands in the lamina propria.
Mucocutaneous Junction and Anal Sacs
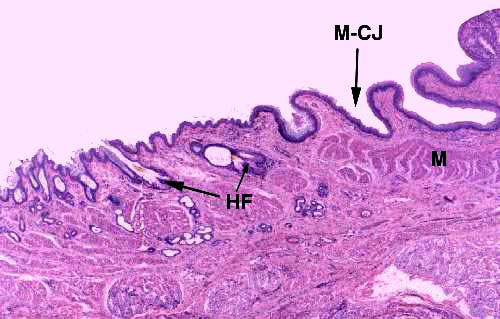 A second "transition" occurs in the anus: it's the mucocutaneous junction at the nether end of the digestive tract (M-CJ). At this point there's no change of epithelial type (it's stratified squamous on both sides) but the post-junctional epithelium is keratinized, as the lining of the anus makes a transition to the general integument.
A second "transition" occurs in the anus: it's the mucocutaneous junction at the nether end of the digestive tract (M-CJ). At this point there's no change of epithelial type (it's stratified squamous on both sides) but the post-junctional epithelium is keratinized, as the lining of the anus makes a transition to the general integument.
To the left in this image you can see typical adnexal structures associated with the integument: a couple of hair follicles (HF) complete with sebaceous glands are visible.
 These sebaceous glands (and some other glands in the region) contribute to the individual's unique scent signature. When one dog meets another for the first time, canine protocol demands that he offer his butt to be sniffed, and that he sniff the other's butt in turn. Dogs who are well-known to each other generally don't do it, but all dogs, when they meet another one for the first time, will do so. Canine courtesy demands that the action be reciprocated. This formality essentially introduces each dog to the other as a specific scent signature, which is then cataloged in memory much as we visually-oriented animals do with faces.
These sebaceous glands (and some other glands in the region) contribute to the individual's unique scent signature. When one dog meets another for the first time, canine protocol demands that he offer his butt to be sniffed, and that he sniff the other's butt in turn. Dogs who are well-known to each other generally don't do it, but all dogs, when they meet another one for the first time, will do so. Canine courtesy demands that the action be reciprocated. This formality essentially introduces each dog to the other as a specific scent signature, which is then cataloged in memory much as we visually-oriented animals do with faces.
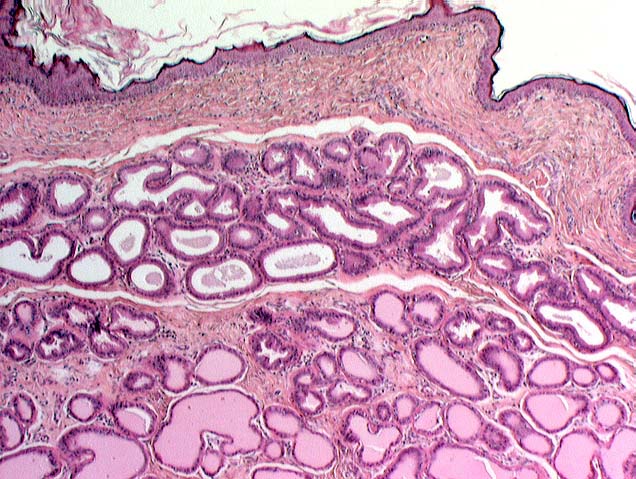 In addition to large sebaceous glands associated with hairs, the anal region of carnivores is the site of the anal
sacs and their associated glands. The breakdown products of these glands, the fecal material, and the secretions of the glands around the anal sacs also provide components of the scent signature. Since in embryological
development both the cranial part of the oral cavity and the caudal
part of the anus originate from depressions in the surface, lined
with surface ectoderm, these sacs and the glands associated with them
are ectodermal derivatives and lined with stratified squamous
epithelium. The glands that surround and empty into the anal sacs are very well-developed in some animals and less so in others.
In addition to large sebaceous glands associated with hairs, the anal region of carnivores is the site of the anal
sacs and their associated glands. The breakdown products of these glands, the fecal material, and the secretions of the glands around the anal sacs also provide components of the scent signature. Since in embryological
development both the cranial part of the oral cavity and the caudal
part of the anus originate from depressions in the surface, lined
with surface ectoderm, these sacs and the glands associated with them
are ectodermal derivatives and lined with stratified squamous
epithelium. The glands that surround and empty into the anal sacs are very well-developed in some animals and less so in others.
In the Mustelidae, the weasels and skunks, the glands are exceptionally well-developed as a defensive mechanism. They're the source of the musky and (in the case of skunks) horribly offensive odor these creatures have. If your dog or cat ever tangles with a skunk, try this remedy for removing the stink.