Objectives for This Exercise
Objectives for This Exercise
VM8054 Veterinary Histology
Author: Dr. Thomas Caceci
PRE-RECORDED LECTURE FOR THIS EXERCISE
 Urinary System Subdivisions
Urinary System SubdivisionsThe urinary system can be subdivided into two major functional subdivisions: the first is the kidney, where the process of manufacture of urine takes place. We'll consider in this exercise the structure of the functional unit of the kidney, the nephron, in detail.
The second subdivision of the system is the excretory passages, all of the structures for collecting urine and draining it out of the body: ureter, the bladder, and urethra. We'll consider these separately, taking the more complicated kidney first.
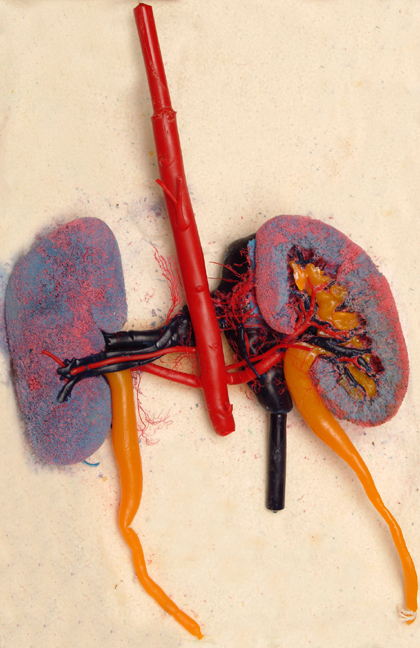
The striking preparation shown at left illustrates the layout of the kidney, its blood supply, and its urinary drainage: this is a corrosion cast preparation, in which plastic has been injected into both the blood vessels and the urinary passages, and then the tissue removed to show these structures in 3 dimensions.
In this preparation, the arterial blood supply has been cast in red plastic; the venous drainage in blue-black. Note that there are red regions in the cortex of the organ: these are locations of the renal corpuscles and their glomeruli (see below). The yellow regions are the renal pelvis, which accepts urine emerging from the lobes of the kidney, and the ureters, which carry it to the urinary bladder.
A preparation like this one clearly emphasizes the extreme vascularity of this organ, and also highlights its role as an interface between the blood compartment and the urine production apparatus. We'll look at the various parts of the system in considerable detail below.
My sincerest thanks to the staff of the VMRCVM's Multidisciplinary Lab and Anatomical Preparation Lab, who supplied this specimen!
The kidney did not evolve primarily as an excretory organ. Though we think of it as one, and though in mammals that's one of its important functions, the primary purpose of the kidney is to control the flux of ions out of the body, and to conserve water. Excretion of nitrogenous wastes evolved as a secondary adaptation, one that took advantage of the kidney's exquisite ability to maintain its internal ionic levels at any desired point.
Capsule of the Kidney
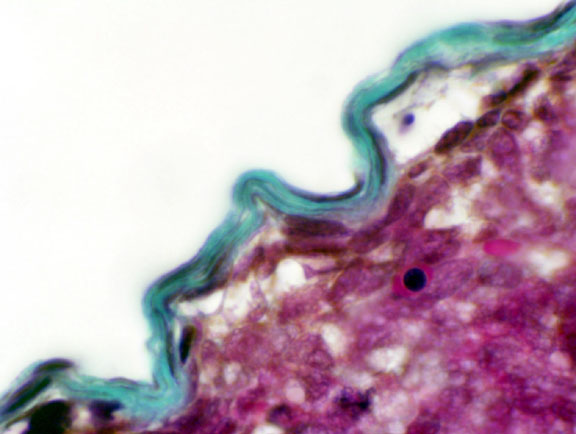 Around the periphery of the kidney is the capsule, a dense collagenous connective tissue envelope. Unlike the capsule of many other organs, this one doesn't send septa down into the parenchyma. There is almost no CT scaffolding in the kidney's internal volume. A scanty amount of CT is present, as a means of ingress and egress for blood vessels, but it's nowhere near as much as would be found in other organs. You will not be able to see it on H&E slides, as the organ has remarkably little fibrous material in it. Slide 152 has been stained with Masson's stain. The capsule of irregular CT is visible as the green-blue covering on the surface of this fetal cat kidney. Various other parts of the kidney are also visible—renal corpuscles and different tubules—but they aren't fully developed in this specimen. We'll examine these in more detail below.
Around the periphery of the kidney is the capsule, a dense collagenous connective tissue envelope. Unlike the capsule of many other organs, this one doesn't send septa down into the parenchyma. There is almost no CT scaffolding in the kidney's internal volume. A scanty amount of CT is present, as a means of ingress and egress for blood vessels, but it's nowhere near as much as would be found in other organs. You will not be able to see it on H&E slides, as the organ has remarkably little fibrous material in it. Slide 152 has been stained with Masson's stain. The capsule of irregular CT is visible as the green-blue covering on the surface of this fetal cat kidney. Various other parts of the kidney are also visible—renal corpuscles and different tubules—but they aren't fully developed in this specimen. We'll examine these in more detail below.
 Cortex, Medulla, and Renal Corpuscles
Cortex, Medulla, and Renal Corpuscles
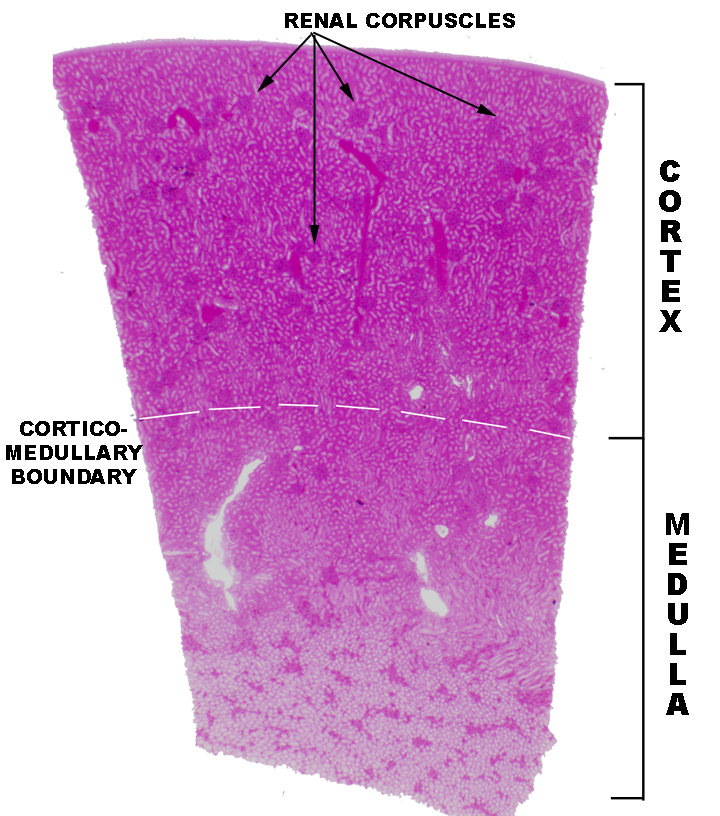
Begin with slide 148. On this slide you can visually differentiate between the cortex and the medulla. The cortex is the outer region, the medulla is the deeper one. The cortex has large numbers of renal corpuscles, discernible as round structures containing small blood vessels. The renal corpuscles are the actual site of blood filtration and the initial stage of urine production. The medulla, deep to the cortex, has no renal corpuscles, only tubules. In the somewhat higher magnification image below, there's a section of the cortex only. Note that although there are many corpuscles in the cortex, the bulk of its substance is composed of the various types of renal tubules that will be discussed below. The medulla has no corpuscles, but the cortex does have tubules.
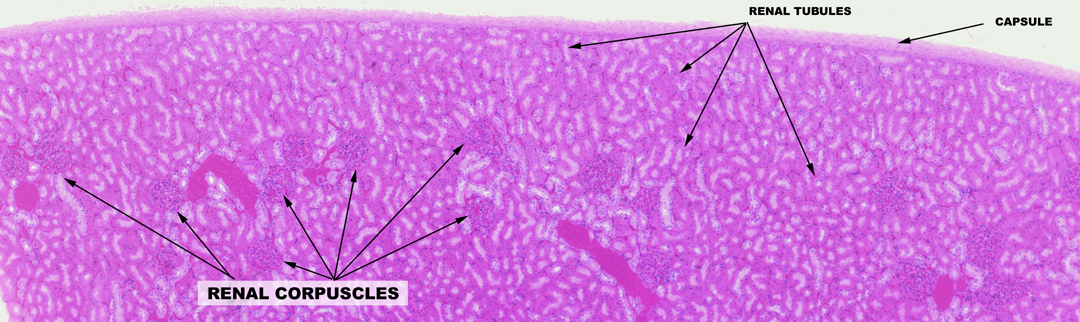
The Renal Corpuscle
The renal corpuscle has two components. The round, hollow renal corpuscle has an outer shell of epithelium, and inside that shell is suspended a capillary complex, the glomerulus. As you can see from the sketch and the actual specimen below, the corpuscle is polarized. One end is the site of entry of blood into the capillary tuft (the vascular pole) and the other end is the beginning of the process of modifying blood filtrate to urine (the urinary pole). The intra-corpuscular space is called Bowman's space. The glomerulus is suspended inside Bowman's space.
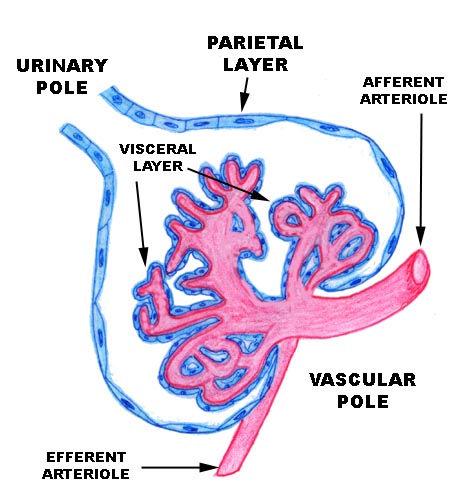

The parietal layer, the outside wall of the corpuscle, is a thin simple squamous type, resting on a basal lamina. It's continuous with the cuboidal epithelium of the proximal tubule at the urinary pole. At the point of entry of the afferent arteriole, the visceral layer is continuous with the parietal layer, sealing off Bowman's space. The corpuscle is thus built like a thin-skinned balloon, in which the glomerulus is suspended. (It's common to hear the entire corpuscle referred to as "the glomerulus," but strictly speaking this is incorrect. The "glomerulus" is just the capillaries.)
The glomerular capillaries are overlain by a specialized visceral layer which consists of the podocytes. The visceral and parietal layer are continuous with each other at the vascular pole. Notice that the urinary pole leads into the first of the nephron's passages, the proximal tubule. So what you have here is a sealed, thin-walled bag around a fluid-filled channel, with a significant pressure across the boundary between the lumen of the glomerular capillaries and Bowman's space.
The Nephron
The nephron is the kidney's basic functional unit. There are a lot of them: about a million nephrons per kidney, so there's a great deal of capacity for processing urine. Enough so that the loss of a kidney to disease or to injury can be compensated by the surviving one!
The nephron has several anatomically distinct regions. These are the renal corpuscle, the proximal tubule, the loop of Henle, and the distal tubule. The basic structure of the corpuscle has been discussed, the rest will be dealt with below.
DISCLAIMER
The nephron is an extraordinarily complicated structure, and comprehending its 3-D architecture of the from a few microscope sections is impossible. Not "difficult": impossible. Serial sections, reconstructions, casting-and-corrosion studies, and other techniques are needed to really appreciate how complex it is. The descriptions in books and lab manuals are the product of years of effort by dozens (if not hundreds) of anatomists. So don't be surprised or frustrated if you find yourself scratching your head as the various parts are described: what you're getting in this exercise is the distillation of all of that effort into a few paragraphs of overwritten prose that can't do its complexity and elegance justice.
Blood Supply
Everything begins at the renal corpuscle. Blood flow determines what happens inside the nephron and outside of it. The kidney is a place in which blood is filtered. The blood flow is the driving force behind urine formation. It's important to consider vascular relationships before we look at the process of urine formation because everything depends on it. Interruption of the blood flow is a major cause of renal failure.
Slide 84 is an "injected" preparation, which will allow you to trace the major blood vessels in the system. In this slide, liquid latex has been pushed through the renal artery under pressure. The latex is thin enough that it flows through the blood vessels. To make the filled vessels more easily visible, the latex has been colored red and set off by a green counterstain.
Blood enters the kidney via the renal artery, and is distributed through a series of smaller vessels. After entering the kidney proper, the large renal artery breaks up into subdivisions. The first such are the interlobar arteries, supplying individual lobes of the organ. It's unlikely that either of these two major vessels will be visible on slide 84, but some small bits of their larger branches will be seen at the hilus. The renal artery and its accompanying renal vein are shown in the corrosion casting above.
From the interlobar arteries, approximately at the corticomedullary junction, several large arcuate arteries (so called because they run along the "arc" of the kidney's curvature) arise. Along their curve they in turn give off smaller branches that run radially to supply the peripheral cortex. You should be able to make out the arcuate arteries as short pieces located at the junction and running parallel to the outer surface of the organ.
The vessels coming off the arcuate arteries are the intralobular arteries, giving rise in turn to the small afferent arterioles, each of which feeds blood into a single glomerulus. One intralobular artery may (and usually will) serve several glomeruli. The small intralobular arteries run radially and as afferent arterioles branch off, they conduct blood into a glomerulus. The glomeruli can be seen "dangling" from afferent arterioles like small berries in this sort of preparation. (Of necessity, this description is actually quite simplified; in most species there are several "orders" of each of the larger arteries, so that one can speak of "primary and secondary arcuates," etc. But the basics are shown well in an injected preparation.)
The arrangement is rather like the water system in a city: large water mains break up into smaller neighborhood lines, which subdivide to serve individual streets, and from the street mains branches come off to each house along the block.
 A single afferent arteriole is dedicated to one specific glomerulus and no other. Flow into the glomerulus leaves it via a second arteriole, the efferent arteriole. This is an important point: blood leaving the glomerulus does not do so in a vein, but in an artery. This unusual situation is needed for two reasons: first, a considerable head of hydrostatic pressure is needed to filter the blood, and venous pressures are not high enough, and; second, the efferent arteriole feeds the capillary bed and venous system that establishes and maintains the kidney's osmotic gradient.
A single afferent arteriole is dedicated to one specific glomerulus and no other. Flow into the glomerulus leaves it via a second arteriole, the efferent arteriole. This is an important point: blood leaving the glomerulus does not do so in a vein, but in an artery. This unusual situation is needed for two reasons: first, a considerable head of hydrostatic pressure is needed to filter the blood, and venous pressures are not high enough, and; second, the efferent arteriole feeds the capillary bed and venous system that establishes and maintains the kidney's osmotic gradient.
The efferent arterioles are somewhat smaller than the afferent ones. Almost immediately upon emerging from the interior of the corpuscle, the efferent arteriole breaks up into the fine capillary networks which surround the loops of the nephrons, the rete mirabile.
Post-Glomerular Blood Flow: Rete Mirabile and Vasa Recta
These post-glomerular capillaries are vital to maintaining a gradient of sodium concentration in the kidney's interstitial spaces. If you were to sample the interstitial fluid at different points, beginning at the deep region of the organ, and moving outward along its radius, you'd find that the concentration of sodium is highest in the deep medulla and lowest in the cortex. That's what's needed for the production of urine hypertonic to the body fluids, as we'll see below.
 The post-glomerular capillaries, as they form, first create the net-like rete mirabile around the entire nephron. The network is most complex close to the glomeruli from which
the capillaries arise; approaching the medulla (i.e., along the deeper portions of the nephron) they form long straight
segments, the vasa recta. These straight regions run very deep, make hairpin bends, and come back up to re-join the rete. Between the sides of each of these loops there are cross-shunts.
The post-glomerular capillaries, as they form, first create the net-like rete mirabile around the entire nephron. The network is most complex close to the glomeruli from which
the capillaries arise; approaching the medulla (i.e., along the deeper portions of the nephron) they form long straight
segments, the vasa recta. These straight regions run very deep, make hairpin bends, and come back up to re-join the rete. Between the sides of each of these loops there are cross-shunts.
Sodium ions are pumped out into the interstitial space of the kidney by active transport, carried out by cells in the wall of the distal tubule (see below). That sodium has to go somewhere, because obviously if it accumulated in the local spaces the concentration would soon get high enough to draw water into the interstitial space by osmosis. That's not good: the water has to stay in the tubules. Where does the excess sodium go? It goes into the vessels of the rete mirabile, i.e., into the blood, by percolating along its natural concentration gradient. If sodium ions aren't needed locally the flow of blood will carry them off into the general circulation.
But...the rete is a tortuous and complicated path. It has numerous cross-shunts at many levels in the vasa recta. By appropriate opening and closing of shunts blood can be channeled anywhere needed, and/or be retained in the immediate vicinity. Since the loops run back up from the deep regions of the medulla to shallow ones, shunting it around within the rete means that the sodium ions aren't carried away at a uniform rate from place to place. By adjusting the flow rate in the various parts of the rete mirabile, the kidney can regulate just how much blood—and hence, how much sodium—is remains localized in different levels of the medulla.
Deep regions need to have a higher concentration than shallower regions, and there has to be a continuous gradient from top to bottom. The rete is crucial to the establishment of this gradient and flow in it is exquisitely regulated to suit conditions almost from minute to minute. Truly a "wonderful net!"
Eventually drainage from all of the post-glomerular capillaries is collected into larger and larger veins until it leaves the kidney via the renal vein, at the hilus. You should be able to make out veins accompanying the larger arteries on slide 84.
We have been talking here about the blood supply, but haven't so far given much consideration to the architecture of the nephron, the basic unit of the kidney, so let's look at it in some detail now.
Urine production is a process of filtration, followed by modification of the filtrate. It's been mentioned that urine starts as an ultrafiltrate of blood (see above) in Bowman's space. All of the formed elements and most of the larger soluble elements (e.g., proteins) have been removed at that point. Propelled by the pressure of continuing filtration, the urine is driven through the rest of the nephron, and the various parts of it modify the product until at the end it's released and drains out through the excretory passages. The exact concentration and composition of the final urine product varies depending on what needs to be cleared and how much water is available to do the job. No better example of the principle that anatomy and physiology are the same thing can be found than the exquisite interrelationships of the nephron and its blood supply.
There are several distinct portions to a nephron. From proximal (i.e. the initial point of filtration into Bowman's space) to distal these are the renal corpuscle, the proximal tubule, the loop of Henle, the distal tubule, and a short collecting tubule. Each nephron is an individual, continuous tubular unit, emptying via its collecting tubule into a collecting duct of larger size, which receives input from many nephrons. A typical mammalian kidney can contain up to a million nephrons.
To continue the analogy above, if the incoming blood supply is laid out like a water supply system, then the outgoing urinary "drainage" is arranged like sewer lines. Each nephron represents one house; its individual drainage line to the street sub-main is via its collecting tubule. These put into the larger main at the junction of two streets, which is analogous to the collecting duct; and these mains then join at the corners into larger and larger mains, until finally the entire output of the neighborhood is channeled into a main flow to the treatment plant.
Now let's look at how the initial filtrate is formed. This takes us back to Bowman's space in the renal corpuscle, and its glomerulus. 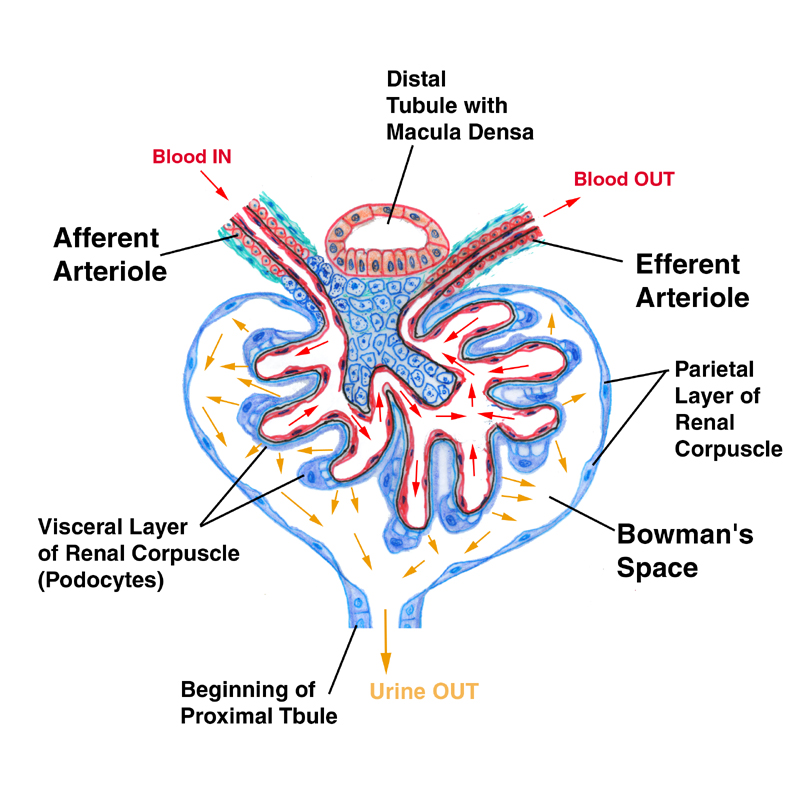 The walls of the capillary tuft are "leaky." Since the blood inside them is under arterial pressure, as it passes through, formed elements are unable to get out but the fluid phase of the blood (the plasma) can do so quite easily. And it does, in large amounts. The capillaries are constantly oozing what is essentially a filtrate of blood plasma. This fills Bowman's space (still under pressure) and is driven into the proximal tubule.
The walls of the capillary tuft are "leaky." Since the blood inside them is under arterial pressure, as it passes through, formed elements are unable to get out but the fluid phase of the blood (the plasma) can do so quite easily. And it does, in large amounts. The capillaries are constantly oozing what is essentially a filtrate of blood plasma. This fills Bowman's space (still under pressure) and is driven into the proximal tubule.
This entire system depends on having a sufficient head of pressure behind the blood to strain it through the barrier of the visceral epithelium and to push it along the tubules. If arterial pressure falls, obviously kidney function will be impaired.
Podocytes
The visceral layer of the renal corpuscle is a very specialized epithelium: it takes the form of podocytes. Podocytes have long, extensively arborized foot processes which wrap around the capillaries of the glomerulus. Podocyte somata can be seen with the light microscope, but their processes can't. The processes cover the entire surface of the capillaries in the glomerulus, except for slits between adjacent processes; these slits are the actual site of filtration. The special nature of this epithelium can only be appreciated in electron micrographs. It forms a "strainer" for the blood.
 You can use this diagram as a key to the electron micrographs below, to clarify the nature of the filtration mechanism and the relationship of podocytes and capillaries.
You can use this diagram as a key to the electron micrographs below, to clarify the nature of the filtration mechanism and the relationship of podocytes and capillaries.
(In this picture only one small section of the wall of the renal corpuscle is shown, with the enclosed loops of the glomerulus' capillaries and their overlying podocyte covering. The area in yellow is Bowman's space, filled with initial filtrate.)
Note that the capillaries of the glomerulus are fenestrated ones, which is what you want in a location like this, where rapid movement of fluid is the goal of the design. The podocytes foot processes lie on top of and surround the capillaries, creating an interlaced meshwork that covers the entire surface.
The mesangial cells shown are intra-corpuscular resident macrophages, which can remove junk that may get through the filtration barrier that shouldn't. (Similar cells called "lacis cells" are found outside the corpuscle, where they perform basic macrophage functions and also serve to seal the system.)
The Basal Lamina and the Nature of the Filtration Barrier
 Now let's look at the structure of the filtration apparatus as it's seen in the transmission EM. In these micrographs you see a glomerular capillary cut in cross section. The fenestrated nature of the endothelium is visible, as is the thick basement membrane outside it. This is combined with the basement membrane of the podocyte processes that are seen as small "islands" of dense arterial spread out along the capillary wall.
Now let's look at the structure of the filtration apparatus as it's seen in the transmission EM. In these micrographs you see a glomerular capillary cut in cross section. The fenestrated nature of the endothelium is visible, as is the thick basement membrane outside it. This is combined with the basement membrane of the podocyte processes that are seen as small "islands" of dense arterial spread out along the capillary wall.
 At somewhat higher magnification (right) the arrangement is more obvious. The podocyte processes are interlaced over the capillary like fingers, with spaces between them. The foot processes are separated from the capillary endothelium by that thick combined basement membrane of the capillary cell and the podocyte itself.
At somewhat higher magnification (right) the arrangement is more obvious. The podocyte processes are interlaced over the capillary like fingers, with spaces between them. The foot processes are separated from the capillary endothelium by that thick combined basement membrane of the capillary cell and the podocyte itself.
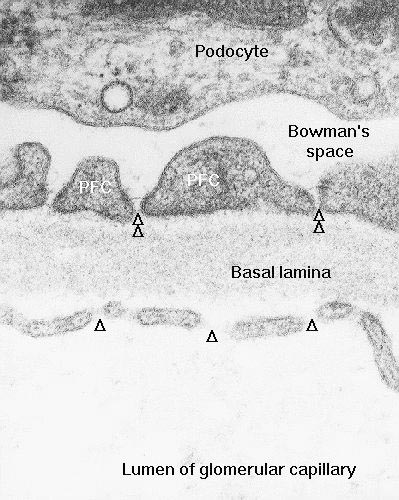 At very high magnification (left) the ends of the foot processes can be seen to be separated by thin slits. Bowman's space is on one side; the capillary is on the other, and the slits are ideally positioned to act as "strainers" for anything that gets out of the blood vessel.
At very high magnification (left) the ends of the foot processes can be seen to be separated by thin slits. Bowman's space is on one side; the capillary is on the other, and the slits are ideally positioned to act as "strainers" for anything that gets out of the blood vessel.
Now, blood passing through the capillaries is under considerable pressure, forcing plasma out of the capillaries and through the slits. Only the liquid phase of the blood gets forced out, and not all of that makes it into Bowman's space: the formed elements are far too large to get through the fenestrations of the capillaries, and the thick combined basement membranes act as microfilters for the larger proteins: nothing much larger than 40 kilodaltons can actually reach Bowman's space under normal conditions. The fluid thus strained into Bowman's space flows thence into the proximal convoluted tubule.
This "sieve" mechanism prevent formed elements of the blood and most large molecules from escaping. Presence of blood or high levels of protein in urine are indicators of damage to the filtration barrier from disease, infection, injury, or parasites.
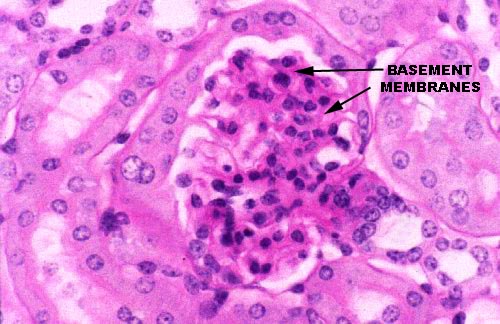 Slide 658 is from a mouse, and has been stained with the PAS reaction. This
makes it possible to see the thick basement membrane in the light microscope as a felt-like layer of amorphous material lying between the podocyte processes and the capillary endothelium. Because there are so many carbohydrates in its chemical composition, it stains heavily with the PAS routine.
Slide 658 is from a mouse, and has been stained with the PAS reaction. This
makes it possible to see the thick basement membrane in the light microscope as a felt-like layer of amorphous material lying between the podocyte processes and the capillary endothelium. Because there are so many carbohydrates in its chemical composition, it stains heavily with the PAS routine.
Proximal Tubule
Most of the cortical tissue around the renal corpuscles is proximal tubules, among which the corpuscles are interspersed. The proximal tubule has a wall of simple cuboidal epithelium with a distinct and prominent microvillous brush border. In microscope preparations the PCT's are often collapsed (due to cessation of glomerular blood flow upon death), but if you look diligently, on slide 148 you should be able to find areas where microvilli are visible. Only the PCT has such a brush border. The other kidney tubules don't.
From Bowman's space, the filtrate passes to the proximal tubule (also called the proximal convoluted tubule, PCT). Here at least 99% of what is present in the filtrate as it leaves Bowman's space is recovered. If it weren't, an animal would die quickly of dehydration and electrolyte loss: on average 1200 liters of blood is filtered per day, but only about a liter of urine is produced. Since small molecules like ions and glucose easily pass through the filtration mechanism, they have to be recovered lest there be a metabolic deficit and/or ionic imbalance.

You can see the brush border in the images above: at left at low magnification and at right at about 1000x. Note the stuff in the lumen of the proximal tubule: that's urine. The PCT's brush border is of course a specialization for increasing surface area for absorptive activity. The PCT extensively modifies the original filtrate. About half a pound of glucose and three pounds of sodium chloride per day are recovered, so that what finally comes out is very different from the starting composition.
 The presence of a brush border is proof that you're looking at a proximal tubule. The brush border in the proximal tubule isn't nearly so distinct as it is in the intestine, but it's nonetheless present, and in favorable sections it can be made out in H&E preparations. The obscurity of the brush border in microscopic sections is due to: 1) the presence of urine in the lumen; and 2) to the collapse of the tubules upon death. Since it's blood pressure that provides the driving force for the filtration mechanism, when the animal dies, the hydrostatic pressure in the tubule drops to zero and it collapses.
The presence of a brush border is proof that you're looking at a proximal tubule. The brush border in the proximal tubule isn't nearly so distinct as it is in the intestine, but it's nonetheless present, and in favorable sections it can be made out in H&E preparations. The obscurity of the brush border in microscopic sections is due to: 1) the presence of urine in the lumen; and 2) to the collapse of the tubules upon death. Since it's blood pressure that provides the driving force for the filtration mechanism, when the animal dies, the hydrostatic pressure in the tubule drops to zero and it collapses.
In the TEM the brush border of the PCT is very well defined (left). The microvilli are similar in structure to those of the intestine, but generally aren't as densely packed or quite so regular. The cells of the PCT's wall are extensively interdigitated along their lateral contact surfaces, as well, to lend mechanical strength to the tubule wall. Since this tubule is a three dimensional structure, you will see microvilli of the brush border cut in all planes, from sagittal to transverse.
The PAS stain makes the PCT's brush border more easily visible. It stains quite heavily using this method, thanks to the presence of a thick glycocalyx. Look for it on slide 689 with this stain.
Loop of Henle
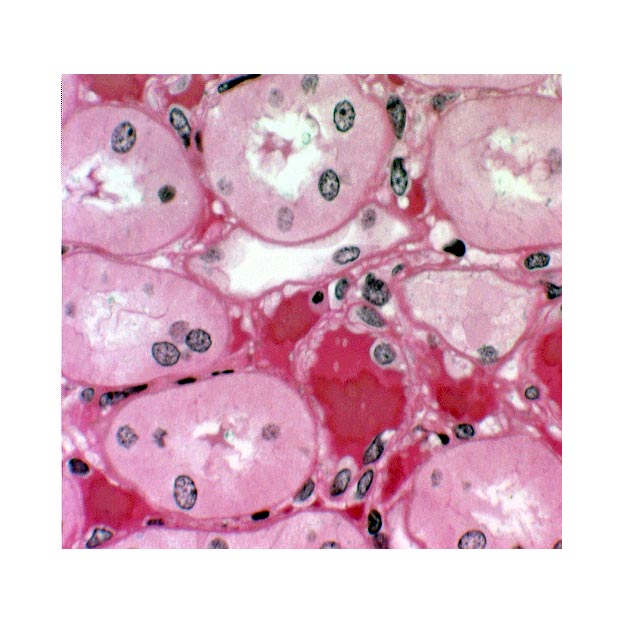 Mammals and birds can make urine that is hypertonic to their body fluids. (Other vertebrates can't.) This
remarkable ability is due to the next portion of the nephron, the loop of Henle, and to the sodium ion gradient through which it passes. The loop of Henle is made of simple squamous epithelium. A loop can be identified on these slides fairly easily by its thin outer wall and empty lumen. If you have a sharp eye and a little luck you'll be able to make out the abrupt transition of the epithelium from cuboidal with microvilli (PCT) to simple squamous (loop). If you're really lucky (or very persistent) you might find the bottom of the loop, where it curves back on its way up to the cortex. Remember that the blood vessels of the rete mirabile and the vasa recta surround these loops.
Mammals and birds can make urine that is hypertonic to their body fluids. (Other vertebrates can't.) This
remarkable ability is due to the next portion of the nephron, the loop of Henle, and to the sodium ion gradient through which it passes. The loop of Henle is made of simple squamous epithelium. A loop can be identified on these slides fairly easily by its thin outer wall and empty lumen. If you have a sharp eye and a little luck you'll be able to make out the abrupt transition of the epithelium from cuboidal with microvilli (PCT) to simple squamous (loop). If you're really lucky (or very persistent) you might find the bottom of the loop, where it curves back on its way up to the cortex. Remember that the blood vessels of the rete mirabile and the vasa recta surround these loops.
This image from slide 148 shows a couple of loops, cut in cross section. The loop of Henle is easy to spot by its wall of squamous epithelium. You can easily tell the thin loop segments from the capillaries of the rete mirabile; the loops are larger and lack erythrocytes.The intimate association of the loops and the blood vessels is obvious, as are the differences in the wall structure between the blood vessels and the loops. The forming urine passes from the last part of the PCT into to the loop.
The loop dives deep into the medullary region of the kidney, forms a hairpin bend, and returns to the level of the cortex again. Filtrate moving through the loop is first carried deep into the medulla; then makes the turn at the deepest part; and returns to the cortex. In so doing it passes through the concentration gradient from the cortex to the medulla and back again.
Distal Tubule
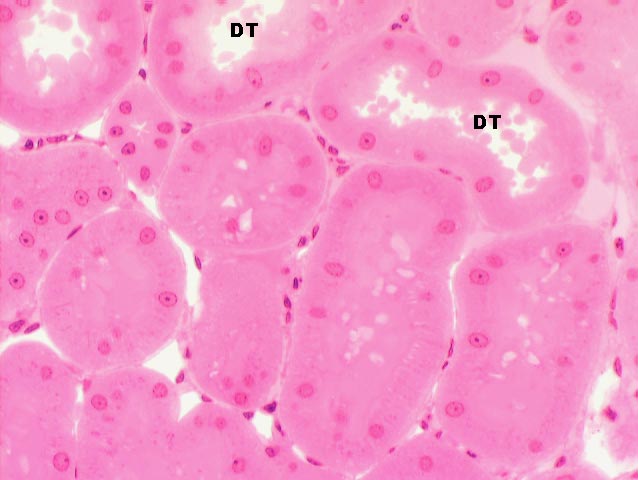 As it comes back up into the cortex region the filtrate passes from the loop of
Henle into the distal tubule (DT). The walls of the DT, like
those of the PCT, are of simple cuboidal epithelium, but they lack a
brush border. They also tend to be larger in cross section than the PCT.
As it comes back up into the cortex region the filtrate passes from the loop of
Henle into the distal tubule (DT). The walls of the DT, like
those of the PCT, are of simple cuboidal epithelium, but they lack a
brush border. They also tend to be larger in cross section than the PCT.
Here's a section with the DT's labeled. There's urine in them. Some of the other tubules in this image are PCT's, but the large open lumen, presence of urine, and most especially lack of a brush border identify these as distal tubules.
The DT isn't specialized for absorption. Its role is to pump sodium ions out of its lumen into the surrounding intracellular space. It does this by active transport, using membrane bound pumps. There's still a fair amount of sodium in the filtrate, and by pumping it into the interstitium, the DT is the source of the "raw material" with which the concentration gradient is established.
 Here's a distal tubule as it's seen in the TEM. In the basal region of the mural cells deep folds enclosing large numbers of mitochondria (M) are visible. The ATP these mitochondria generate is used to drive the active transport pumps that move sodium out of the distal tubule and into the interstitial space of the kidney. It's the transport of these ions (coupled with their selective removal via the rete mirabile) that establishes the osmotic gradient necessary for the kidney's function of making hypertonic urine. Active transport is required because the concentration in the interstitium may be very high, much higher than in the tubule, so the sodium has to be pumped out.
Here's a distal tubule as it's seen in the TEM. In the basal region of the mural cells deep folds enclosing large numbers of mitochondria (M) are visible. The ATP these mitochondria generate is used to drive the active transport pumps that move sodium out of the distal tubule and into the interstitial space of the kidney. It's the transport of these ions (coupled with their selective removal via the rete mirabile) that establishes the osmotic gradient necessary for the kidney's function of making hypertonic urine. Active transport is required because the concentration in the interstitium may be very high, much higher than in the tubule, so the sodium has to be pumped out.
The cells are "sealed" at the top with an occluding junction. Occluding junctions are impermeable even to ions, and their presence implies that there's a good reason to prevent free percolation of materials from one side of the junction to the other. Control of ion passage is vital, and the only way those ions are going to be allowed to get out of the tubule is by a controlled active transport. Notice also the complete absence of microvilli on the free surface. This is a clear indication that absorption from and/or secretion into the lumen isn't a function of these cells.
In cross section the DT's are usually quite a bit larger than PCT's, and will show 4 to 6 nuclei, rather than the 2 to 4 seen in the PCT. Also, the nuclei of DT cells tend to be closer to the lumen, and to bulge out into it somewhat.
Obviously, since the DT and the post-glomerular blood flow are so intimately integrated, there must be some sort of "feedback" between them. There's one special region in the wall of the DT which is probably involved in this coordination. Locating it will definitively identify the distal tubule.
As it comes back up to the cortical level, a portion of the DT comes to lie in close proximity to the entry point of the afferent arteriole into the renal corpuscle: this association is always between a DCT and its own corpuscle. There forms at this point a peculiar association of cells of the tubule, the arteriolar wall, and the nearby kidney parenchyma, called the juxtaglomerular apparatus. The juxtaglomerular apparatus is involved in the regulation of systemic blood pressure, and hence indirectly controls glomerular blood flow.
 In the wall of the DT at the point where the tubule and the afferent arteriole
are in contact cells are taller than the normal and much more crowded together.
You'll have little trouble spotting this structure on slide 148. This is
the macula densa (Latin for "dense spot"). Look for an area
near an afferent arteriole, in which nuclei are crammed tightly; there may be
as many as 10, distinctly polarized towards the arteriole where it enters the
capsule. If you were standing inside the distal tubule, you'd see it as such. It's as if there were more bricks in a certain part of a wall than in other parts.
In the wall of the DT at the point where the tubule and the afferent arteriole
are in contact cells are taller than the normal and much more crowded together.
You'll have little trouble spotting this structure on slide 148. This is
the macula densa (Latin for "dense spot"). Look for an area
near an afferent arteriole, in which nuclei are crammed tightly; there may be
as many as 10, distinctly polarized towards the arteriole where it enters the
capsule. If you were standing inside the distal tubule, you'd see it as such. It's as if there were more bricks in a certain part of a wall than in other parts.
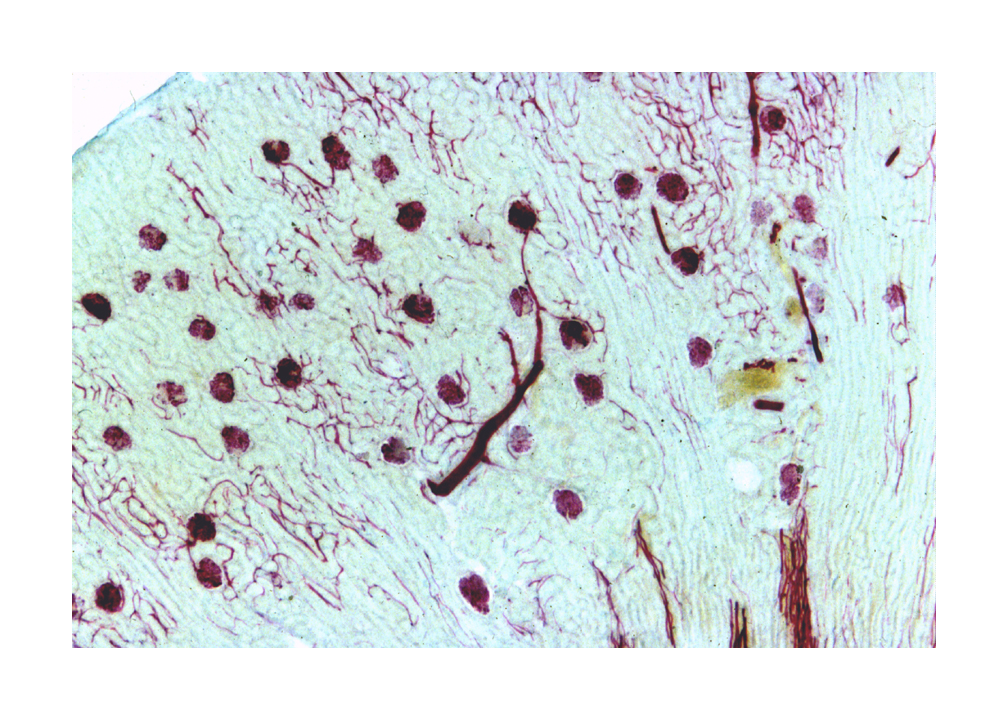
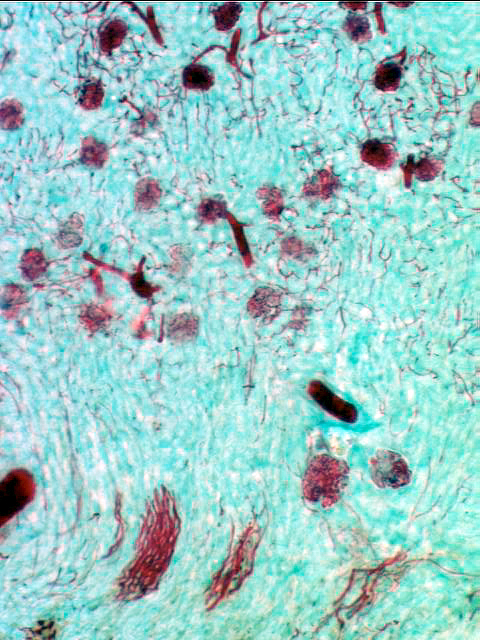
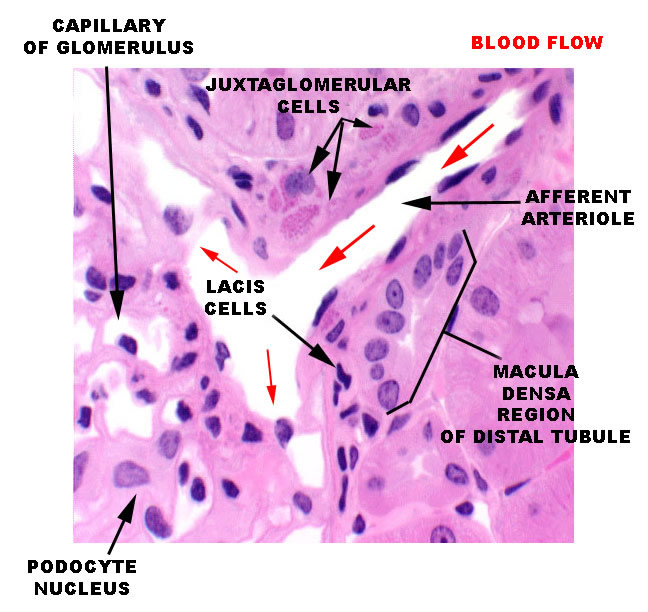 The MD is a very localized structure. This specific and structurally distinct region is always associated with the afferent arteriole of its own nephron. The glomerulus you see immediately to the right of the macula densa is the same one whose filtrate will eventually pass through that DT. The macula densa is polarized away from the lumen of the tubule, and towards the adjacent specialized cells in the wall of the afferent arteriole. It's thought to be a sensory structure involved in the release of renin from another portion of the juxtaglomerular apparatus, and thus indirectly involved in controlling of the flow through its nephron and others. The image at left is from the vascular pole of a renal corpuscle: the afferent arteriole is bringing blood in at upper right and it's being distributed through the glomerular capillaries. The region of the arteriole close up to the wall of the corpuscle has several cells in it that contain pink-stained granules of renin (see below). these are the juxtaglomerular cells. In close association with these specialized cells, the macula densa is visible. Between the MD and the nearest Juxtaglomerular cells are a few lacis cells of uncertain function.
The MD is a very localized structure. This specific and structurally distinct region is always associated with the afferent arteriole of its own nephron. The glomerulus you see immediately to the right of the macula densa is the same one whose filtrate will eventually pass through that DT. The macula densa is polarized away from the lumen of the tubule, and towards the adjacent specialized cells in the wall of the afferent arteriole. It's thought to be a sensory structure involved in the release of renin from another portion of the juxtaglomerular apparatus, and thus indirectly involved in controlling of the flow through its nephron and others. The image at left is from the vascular pole of a renal corpuscle: the afferent arteriole is bringing blood in at upper right and it's being distributed through the glomerular capillaries. The region of the arteriole close up to the wall of the corpuscle has several cells in it that contain pink-stained granules of renin (see below). these are the juxtaglomerular cells. In close association with these specialized cells, the macula densa is visible. Between the MD and the nearest Juxtaglomerular cells are a few lacis cells of uncertain function.
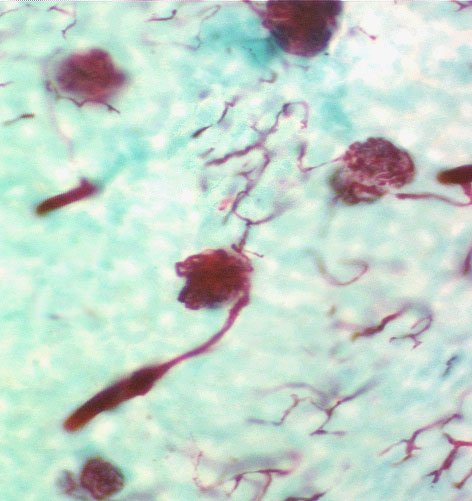
The macula densa is only one part of the complete juxtaglomerular apparatus. The specialized juxtaglomerular cells that form a a "sleeve" around the lumen of the arteriole are part of it, too. These cells are modified smooth muscle cells, easily recognized on slide 148 by the presence of bright pink renin granules in their cytoplasm. Renin is a hormone involved in regulation of blood pressure. The juxtaglomerular cells in the afferent arteriole are thought to be specialized blood pressure receptors, with the capacity to release renin when needed.
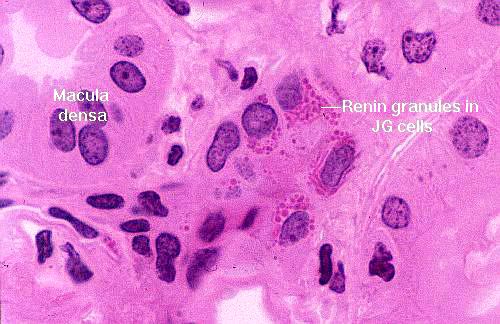 In this image the granules are very noticeable: they're polarized towards the blood vessel, because as it true of all hormones, renin is secreted into the blood.
In this image the granules are very noticeable: they're polarized towards the blood vessel, because as it true of all hormones, renin is secreted into the blood.
A fall in systemic blood pressure triggers a response in the juxtaglomerular cells, which causes renin release into the blood. The macula densa cells may function as chemoreceptors, sensing a drop in sodium concentration of the DT, to trigger renin release. The intimate association of the MD and its own blood supply enables it to monitor and respond to these changes directly and immediately. Via a series of enzymatic reactions, renin indirectly causes peripheral vasoconstriction and increased water uptake in the kidney, to minimize fluid loss and pressure drop.
Renin is a protease. It acts on a circulating plasma protein, angiotensinogen, to cleave off a segment and convert this precursor material into angiotensin I. This, in turn, is cleaved further by a protease in the lung to produce the hormone angiotensin II. Angiotensin II is a powerful vasoconstrictor, that raises blood pressure and indirectly influences renal flow rates.
Renin also probably has a more direct local effect on flow through the nephron. Recent evidence shows that the juxtaglomerular cells have all the necessary enzyme pathways to produce angiotensin themselves using this renin-angiotensin I-angiotensin II cascade. Thus, it's possible that in addition to a general mediation of systemic blood pressure, the juxtaglomerular apparatus can release active angiotensin II into the local area, and regulate glomerular blood flow by inducing localized vasoconstriction. The macula densa is thought to be a detector for local salt concentrations and flow rate, which signals other elements of the juxtaglomerular apparatus to adjust the flow rate as needed.
Collecting Tubules and Collecting Ducts
Strictly speaking, the end of the DT marks the terminus of the nephron, but even when the urine-to-be reaches that point, its modification isn't finished. The last step is to pass it through the concentration gradient a second time, from cortex to medulla, as a way to make its final concentration correct. This happens in the collecting tubules and collecting ducts.

 The collecting tubule receives the filtrate from the DT and carries it on its
last—and most important— trip down through the medulla and out to the world
beyond. Collecting tubules are quite large, with walls formed of cuboidal
epithelium. They're larger than the DT and the nuclei of their mural cells
bulge somewhat into the lumen. The cells themselves are rather pale staining
compared to those of the other tubules. Large collecting ducts are formed by
the fusion of a number of collecting tubules, and drain several nephrons.
The collecting tubule receives the filtrate from the DT and carries it on its
last—and most important— trip down through the medulla and out to the world
beyond. Collecting tubules are quite large, with walls formed of cuboidal
epithelium. They're larger than the DT and the nuclei of their mural cells
bulge somewhat into the lumen. The cells themselves are rather pale staining
compared to those of the other tubules. Large collecting ducts are formed by
the fusion of a number of collecting tubules, and drain several nephrons.
In the image at left, the difference in appearance of the collecting ducts is pretty clear. This is a pretty typical field from the renal medulla: the loops of Henle (TLH) descend quite a ways into the medulla, and the collecting ducts pass all the way through it. Thus a plane of a section through the medulla will intersect several different types of tubule wall structure. The collecting duct (CD) is the largest of these, and it has a wall of distinctly high cuboidal to columnar epithelium. After the Loop has made its hairpin turn, the ascending part of the distal tubule (DT) begins. You can almost always tell the PCT and DT apart in the medullary region by their size (the DT is larger in diameter) and the fact that in a PCT, typically only one or two nuclei of the wall cells will be visible; more than that are usually seen in a DT profile. Capillaries of the vasa recta (Cap) are also visible.
Recall the mechanical analogy of the sewer line: input to the sewer line comes from individual toilets (nephrons) brought there by the separate house drain lines (the collecting tubules). Each drain line joins the sewer main running down the middle of the street (the collecting duct). The large ducts open out onto the tip of the renal pyramid, or on the renal crest.
The epithelium of the collecting ducts is sensitive to antidiuretic hormone (ADH). When ADH levels are high, the walls become very permeable to water. When ADH levels are low, the opposite occurs; duct walls are impermeable to water. Hormonal control of duct permeability adjusts the final volume and concentration of urine voided.
The collecting ducts pass through the entire length of the intra-kidney ionic gradient. As forming urine makes the final trip from the cortex through the medulla, it passes through regions in which there is greater and greater concentration of sodium ions in the extracellular spaces. If the ADH levels are low, and the duct is permeable water is drawn out in a futile attempt to equilibrate this space. The result is a small volume of concentrated urine. If ADH levels are high, water is retained in the collecting duct, and the result is a copious dilute urine. Thus it is in the collecting ducts and tubules that the final concentration is determined.
Having produced urine, it's necessary to get rid of it. As mentioned, it drips from the tips of the renal pyramids into the renal pelvis (or the renal crest in some species) into the upper end of the collecting system, and flows through a series of tubes to the outside.
 The renal pelvis is the upper end of the ureter, attached just below the tips of
the kidney's lobes. It catches the urine and drains it out. It's a CT
"cup" lined with urinary epithelium. On slide 152 you can see the tip of the lobe, whence drips out the final urine. This is the tip of the renal crest from a preparation of fetal cat kidney. The openings you see in it are the ends of the large collecting ducts (D), each draining a large number of nephrons. These ducts are lined with a high cuboidal to columnar epithelium and urine drips from them in a more or less continuous stream into the space between the renal pelvis (RP) and the renal crest.Some of the urine has been coagulated and trapped in the specimen (Ur).
The renal pelvis is the upper end of the ureter, attached just below the tips of
the kidney's lobes. It catches the urine and drains it out. It's a CT
"cup" lined with urinary epithelium. On slide 152 you can see the tip of the lobe, whence drips out the final urine. This is the tip of the renal crest from a preparation of fetal cat kidney. The openings you see in it are the ends of the large collecting ducts (D), each draining a large number of nephrons. These ducts are lined with a high cuboidal to columnar epithelium and urine drips from them in a more or less continuous stream into the space between the renal pelvis (RP) and the renal crest.Some of the urine has been coagulated and trapped in the specimen (Ur).
Cats (as do other carnivores) have developed a fusion of the ends of the renal lobes to form a single renal crest, rather than having each lobe empty separately into its own "funnel" as in other species (see the corrosion specimen at the beginning of the exercise for the alternative arrangement). The upper end of the pelvis is seen at the bottom of the field, stained green-blue. The lower end of this funnel shaped structure is attached to the ureter, and the upper edges are sealed off with CT to prevent leakage of urine. Regardless of the gross anatomy, urine produced in the nephrons is moving through them under pressure and it has to go somewhere, out the open ends of the ducts.
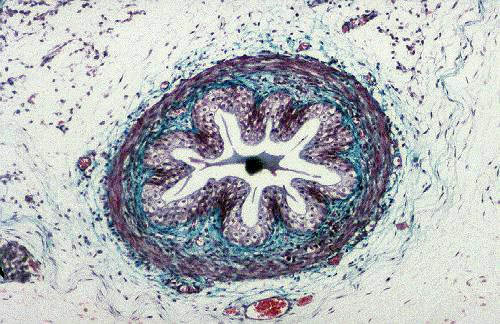
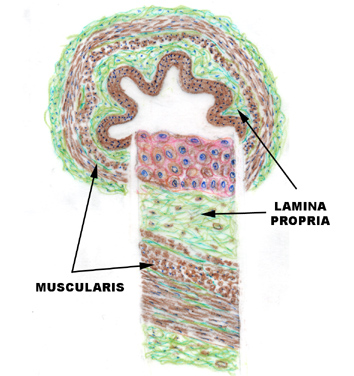 The ureter at left is from slide 152, the fetal cat preparation stained with Masson's method. The wall of the ureter at this level has a thin muscular outer tunic, a CT lamina propria, and a lining of urinary epithelium. These features are marked in the sketch for reference to the actual specimen. Slide 657 is the midlevel of the ureter of a dog. In this longitudinal section,
the nature of the epithelium is quite evident; and so is the muscularity of the
wall. The epithelium rests on a basal lamina, and it has a support of CT in the
form of a lamina propria/submucosa made of collagenous fibers. It's a really nice example; the epithelial structure is unmistakable, and the
muscular wall very thick. You can see in these cross sections that there are two
layers of muscle in the wall: an inner longitudinal one, and an outer circular
one. The extensive folding of the epithelium results from the tonus of the
muscle around it. When urine passes through, the lumen of the tube dilates
under the pressure.
The ureter at left is from slide 152, the fetal cat preparation stained with Masson's method. The wall of the ureter at this level has a thin muscular outer tunic, a CT lamina propria, and a lining of urinary epithelium. These features are marked in the sketch for reference to the actual specimen. Slide 657 is the midlevel of the ureter of a dog. In this longitudinal section,
the nature of the epithelium is quite evident; and so is the muscularity of the
wall. The epithelium rests on a basal lamina, and it has a support of CT in the
form of a lamina propria/submucosa made of collagenous fibers. It's a really nice example; the epithelial structure is unmistakable, and the
muscular wall very thick. You can see in these cross sections that there are two
layers of muscle in the wall: an inner longitudinal one, and an outer circular
one. The extensive folding of the epithelium results from the tonus of the
muscle around it. When urine passes through, the lumen of the tube dilates
under the pressure.
Urinary Bladder
Slide 723 is the urinary bladder. The bladder is a temporary storage organ for urine, whose capacity (in humans) can be measured in terms of milliliters, quarts, or in miles between rest stops on the Interstate highways.
The wall of the bladder is similar to that of the ureter, but there are said to be three layers of smooth muscle: inner and outer layers are longitudinal, and the middle more or less circular. The orientation of these is less than perfect, and given the shape of the bladder, this is perhaps to be expected. There is also a fair amount of elastic connective tissue in the wall to provide for elasticity and distensibility.
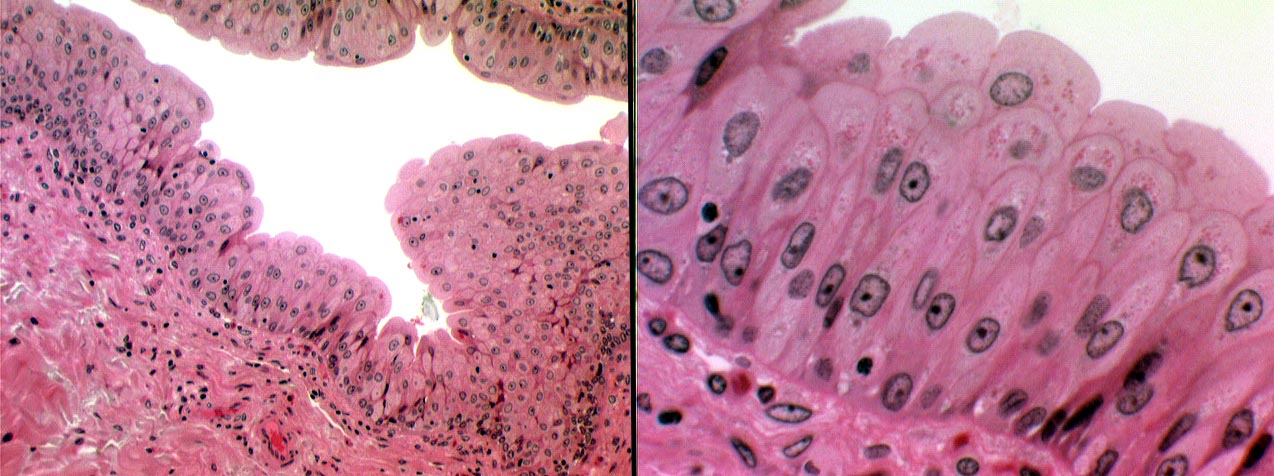
The lining is urinary epithelium, extensively folded and puckered in the relaxed state. Refer to Exercise 4 for a discussion of this special epithelial type, and to the image above. The scalloped free border is visible, and the inclusion bodies that are characteristic of the urinary epithelial cells are evident. These are actually areas of spare plasma membrane. When the bladder is relaxed (as it is in these images) the excess membrane is "tucked in" but remains attached to the surface; when the bladder expands, there is ample membrane to prevent the surface from being ruptured.
 Urethra
Urethra
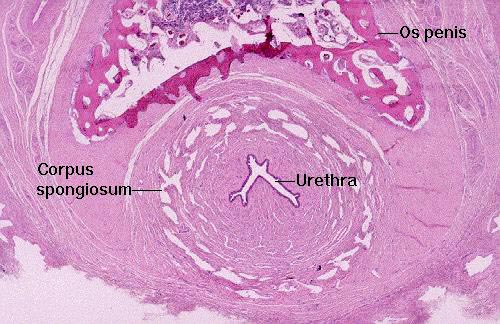
Slide 707 has on it a cross section of a canine penis, and in the center you will see a section of the final passageway, the urethra, which in males does dual duty as part of the urinary tract and the reproductive system. The end of penile urethra has a lining of stratified squamous epithelium; farther up in the pelvic urethra, nearer the bladder, the lining is still urinary epithelium.
In this cross section of a dog penis (distal end) you see the urethra in the middle. Around it is the corpus spongiosum, a series of blood chambers that keep the lumen patent under conditions of tumescence. The urethra in males serves as a dual purpose route for the expulsion of sperm and urine. In the dog, as in other species (but not humans) there's a bone, the os penis, on the dorsal side.