Connective Tissues Proper
A threefold cord is not quickly broken.
—Ecclesiastes III:12
Objectives for
this Exercise
CASE VIGNETTE 1
CASE VIGNETTE 2
SLIDES USED IN THIS EXERCISE: 29, 32, UWO120, 11, 12, 35, 1210, 33, UC008, 113, 27, 40, 1193
THERE ARE PRE-RECORDED LECTURES FOR THIS EXERCISE
Please go to the listing page for these to download and view them
Cells of Connective Tissues
Fibrous Connective Tissues
Special Connective Tissue
"Proper" versus "Special" CT's
Connective tissues as a group are those having protective or supportive
functions. All CT's have three elements: fibers, cells, and non-cellular
matrix material. The proportions of the three vary widely in different
CT's, and it's the proportions among them that give any given CT its
characteristic properties.The term connective tissue proper is used for those examples in which the
fibrous component is the predominant feature. It's the group which is most
often spoken of as "connective tissue," although really it's only one
of several subclassifications.
Special connective tissues are a heterogeneous group. They're found in
specific places, they have specific functions, and they're not nearly as common
as the "proper" CT in terms of distribution. This term is applied to reticular,
mucous, elastic, and adipose connective tissue.
Fibers of Proper CT's
 In most CT proper, the predominant fiber type is collagen. Collagen is
far and away the most typical fiber of any CT's, and in fact, collectively collagen(s) is (are) the most abundant protein(s) in the body. If you somehow removed everything from an animal except its collagen, you could still identify all the organs and parts of organs grossly.
In most CT proper, the predominant fiber type is collagen. Collagen is
far and away the most typical fiber of any CT's, and in fact, collectively collagen(s) is (are) the most abundant protein(s) in the body. If you somehow removed everything from an animal except its collagen, you could still identify all the organs and parts of organs grossly.
Actually, collagen isn't a single
material; depending on what reference you consult, there are at least six, and
perhaps as many as 15 chemically distinct collagen variants. Some are found in specific locations, but for our purposes,
we can lump them all together as "collagen" and note the
differences where appropriate. Morphologically almost all types of collagen are
indistinguishable at the LM level. Collagen has been likened to a "biological rope."
It's composed of
smaller subunits (tropocollagen molecules) laid together in triplets,
somewhat like braids. This produces immensely strong fibrils, which are then
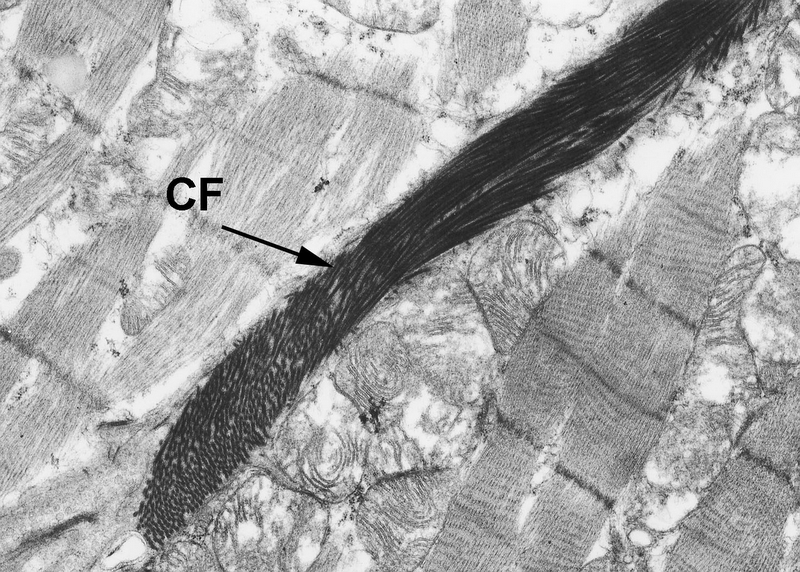
bound together to form larger fibers when necessary. The electron microscopic image at left shows the typical appearance of collagen fibrils.
The example above at left is from the muscle of the heart, and the collagen fibrils are dense single strands, forming a ribbon-like fiber that lies between the muscle bundles. In longitudinal section typical collagen has a "banded" appearance, as seen at right. Note also the varying fibril sizes. Chemically, collagen is
characterized by a high proportion of hydroxyproline, an amino acid
derivative more or less unique to this material. The content of hydroxyproline
in a tissue is considered a measure of its collagen content.
Cells of Connective Tissues: "Fibroblasts" and "Fibrocytes"
 The cells of most CT's, regardless of whether they are "proper" or
"special" types, are derived from the same primordial line, and hence
there is considerable overlap in the cell types found in different CT's. If you
examine fibrous CT's closely you'll be able to see the cells which make and
maintain the fibers. These are the fibroblasts and fibrocytes,
the most numerous and important of the cell types of most CT's. We'll consider
other cell types later, but for now let's discuss these two in some more detail.
The cells of most CT's, regardless of whether they are "proper" or
"special" types, are derived from the same primordial line, and hence
there is considerable overlap in the cell types found in different CT's. If you
examine fibrous CT's closely you'll be able to see the cells which make and
maintain the fibers. These are the fibroblasts and fibrocytes,
the most numerous and important of the cell types of most CT's. We'll consider
other cell types later, but for now let's discuss these two in some more detail.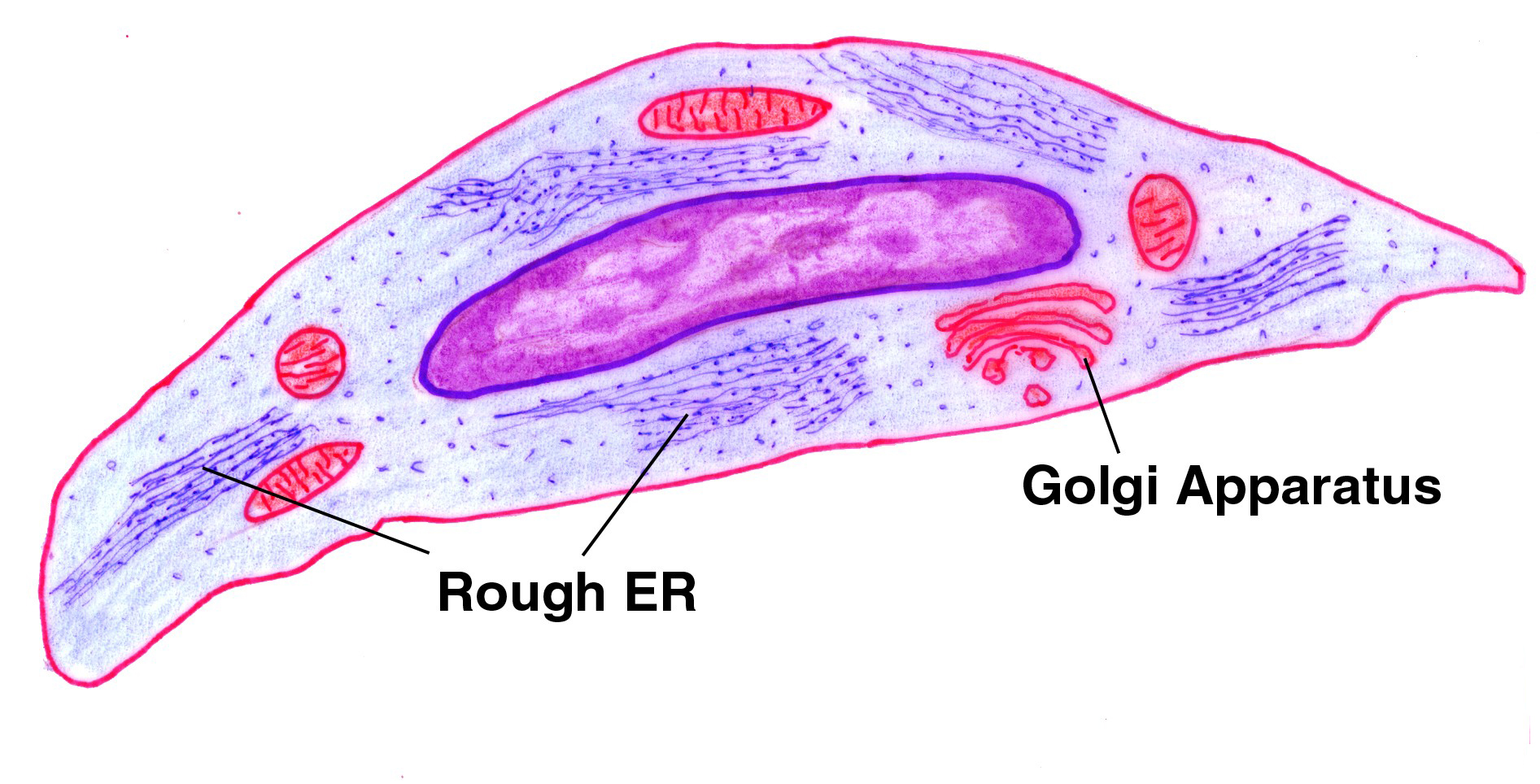
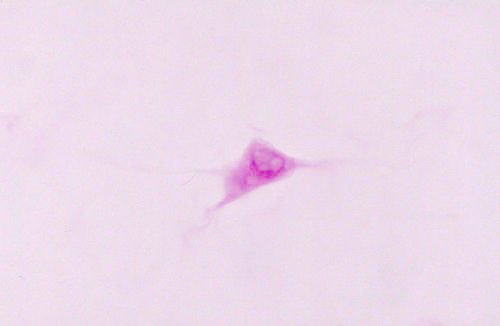
This field (above left) is of a single fibroblast, the Protean cell that makes all the components of CT's, and whose stem cell line also produces other CT cells. (The same cell line that generates fibroblasts produces the osteoblasts of bone, the adipocytes of fat , and the chondrocytes in cartilage). The fibroblast is a seen here en face, that is, looking down on it from the top.
The sketch shows a little more clearly the nature of the cell. Fibroblasts are flattened and like this one, usually have a spiky and irregular outline. They're sometimes referred to as "stellate" in shape (from Latin, stella = "star"). You'll note that there are some wispy extensions of the cell's cytoplasm: these "arms" are sitting on collagen fibers (which are invisible in this image) and acting to maintain and repair those fibers. 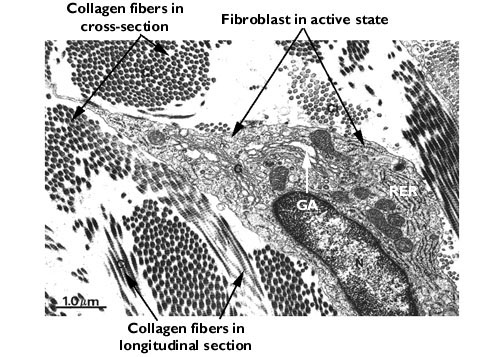 The importance of fibroblasts can't be overestimated. They're involved in normal growth, healing, wound repair, and the day-to-day physiological activities of every tissue and organ in the body. The fibroblast does everything.
The importance of fibroblasts can't be overestimated. They're involved in normal growth, healing, wound repair, and the day-to-day physiological activities of every tissue and organ in the body. The fibroblast does everything.
Fibroblasts are thought to be able to de-differentiate back to earlier stages in their development and then re-differentiate into some other cell type. For example, they can regress back, shift gears, and re-differentiate into osteoblasts, to aid in the production of bone; into chondrocytes under some circumstances; or even, if need be, into adipose cells.
It's the fibroblasts of the intermuscular CT that form the scar tissue when muscle cells are destroyed, weaving new collagen fibers to fill the gaps left 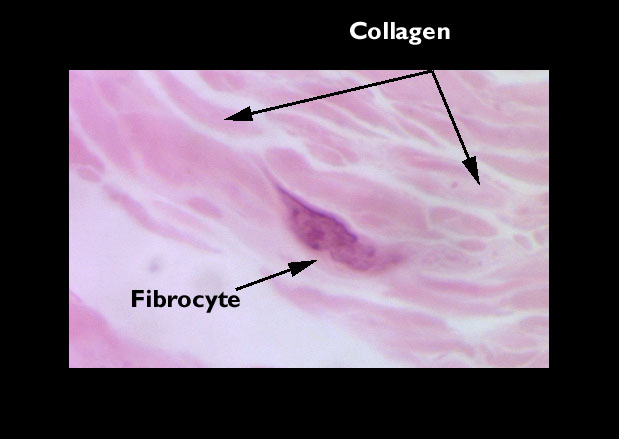 by dead myocytes. This is the process of fibrosis, a vitally important concept in pathology. Differentiated as chondroblasts (the stem cells of cartilage) they make not only the collagen and/or elastic fibers appropriate for the cartilage type, they make the matrix as well. In the light microscope, there isn't much to be seen without special stains, but (as in the micrograph shown here) it's usually possible to make out the nucleus and the boundaries of the cell. Special stains that emphasize the acidic nature of the cytoplasm make it easier to define them in light micrographs. In the EM, fibroblasts are recognizable by their shape, their large complement of RER, and their large, prominent nucleolus when they're actively making materials (above, right).
by dead myocytes. This is the process of fibrosis, a vitally important concept in pathology. Differentiated as chondroblasts (the stem cells of cartilage) they make not only the collagen and/or elastic fibers appropriate for the cartilage type, they make the matrix as well. In the light microscope, there isn't much to be seen without special stains, but (as in the micrograph shown here) it's usually possible to make out the nucleus and the boundaries of the cell. Special stains that emphasize the acidic nature of the cytoplasm make it easier to define them in light micrographs. In the EM, fibroblasts are recognizable by their shape, their large complement of RER, and their large, prominent nucleolus when they're actively making materials (above, right).
Interference with their activities causes a vast array of clinical problems. Scurvy, the disease brought on by lack of vitamin C in humans, is an example. The deficiency affects a specific pathway in collagen synthesis, for which vitamin C is required. Dietary deficiency of vitamin C has consequences ranging from skin sores, anemia, edema, ulcerated gums, loosened teeth, and hemorrhage of mucous membranes. Restoration of the vitamin allows the fibroblasts to work normally and cures the problem.
Some authorities make a distinction between "fibroblasts" and
"fibrocytes." In this nomenclature a "fibroblast" is a cell actively producing
collagen fibers and (usually) matrix material as well. It has the distinctive characteristic of protein secreting cells: a vesicular
nucleus, comparatively basophilic cytoplasm, and prominent nucleoli. The term "fibrocyte" refers to cells which have passed into a quiescent phase,
in which they're concerned with fiber maintenance, not production.
In wound repair and normal growth, active fibroblasts are quite common but their
proportion of the total CT cell population is reduced when things are okay and
there isn't much work to be done other than keeping the fibers in good shape.
The distinction between "-blast" and "-cyte" is largely
academic from the histologist's point of view, since the change from one state
to another is reversible; and so throughout these exercises, for the sake of
simplicity I'll just refer to them all as "fibroblasts" and be done
with it.
The fibroblasts are very thin and flattened, and lie atop the fibers they've
made. Since there's so little cytoplasm, you generally won't see it, as it
doesn't take up much stain. Fibroblasts are usually seen as spindle-shaped
nuclei lying along the fibers when you look at them "on edge," but if
you view one en face (i.e. from the "top" as if you
were hovering over it) you'll see an oval nucleus and some wispy cytoplasm
around it, usually stellate in shape. If the cell is active, you may detect
some basophilia.
It's much easier to find these cells and see what they look
like in a loose collagenous CT than in a dense one (see below). Note that
fibroblasts lie between and on top of their fibers, not inside them! Collagen
is an extracellular, insoluble material. Although its precursors are manufactured
in cells, these are extruded and "cured" extracellularly to form the
final fiber. Hence the cells are not embedded inside the fibers.
Collagenous CT's
Irregular CT's
Loose or areolar CT is fairly common, and can be subclassified as "regular" or "irregular" depending on the arrangements of
its fibers. Loose irregular CT is found in the lamina propria of the
digestive tract, and is so heavily cellular that the fibers are not usually
visible. Dense irregular CT is more easily demonstrated.
These two examples are of dense irregular collagenous CT from slide 7, skin from a pig. In the image below left, the dark purple U-shaped band at the top of the field is the epithelium of the skin, and the rest is the CT. Immediately below the surface epithelium there's a broad area of pink-staining material, composed of collagen fibers making up the mesh of dense irregular CT that supports the epithelium. 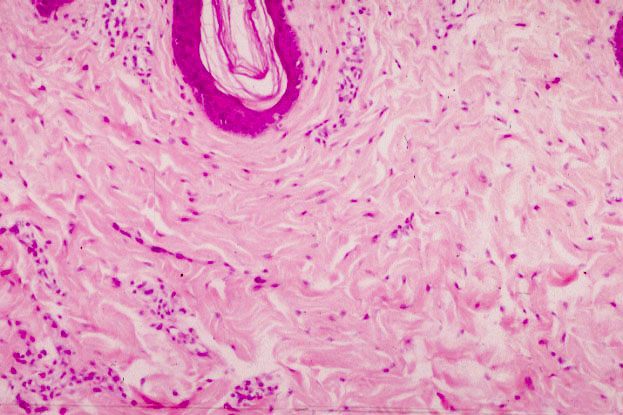 In this type of connective tissue, the fibers (principally collagen type I, but with a few other fiber types mixed in) run in all directions, and have no particular orientation. Connective tissue of this type usually has blood vessels running through it in histological sections. A small artery is visible in the higher-power image at right.
In this type of connective tissue, the fibers (principally collagen type I, but with a few other fiber types mixed in) run in all directions, and have no particular orientation. Connective tissue of this type usually has blood vessels running through it in histological sections. A small artery is visible in the higher-power image at right.
Dense irregular CT is usually covered by some sort of epithelium. Epithelium is avascular, so the CT is the route by which nutrient and waste transport services are provided. You are likely to see blood vessels in almost any CT specimen you examine, certainly in the irregular collagenous types. The little purple dots scattered through the field are the nuclei of fibroblasts/fibrocytes seated on the fibers to maintain and repair them. (Keep in mind that the actual fibers are extracellular structures. The fibroblasts extrude soluble precursors and link them together into the insoluble form outside the cell.)
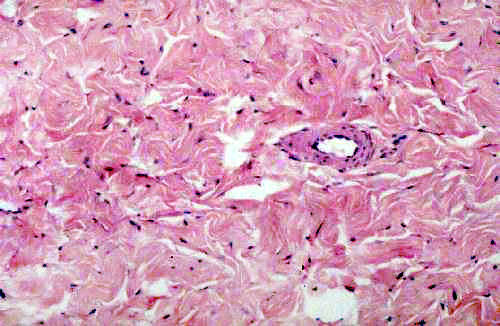 Most CT's are derived from embryonic mesoderm, as are blood vessels (and for that matter, the blood itself, which is yet another form of CT). Hence it shouldn't be surprising that blood vessels are found in such locations.
Most CT's are derived from embryonic mesoderm, as are blood vessels (and for that matter, the blood itself, which is yet another form of CT). Hence it shouldn't be surprising that blood vessels are found in such locations.
The fibers of an irregular CT are randomly arranged. In the region
immediately below the surface of the skin the collagen fibers are fairly
densely packed, hence the description of this tissue as "dense
irregular." Deeper into the sample (i.e., farther from the surface)
you will see the spaces between the fibers become larger. This is a "loose
irregular" arrangement. A mechanical analogy of the difference between
dense and loose irregular CT is the difference between hat felt (dense
irregular) and the wad of lint you pull out of your clothes dryer's filter
screen (loose irregular). Both have randomly arranged fibers, but one is more
tightly compressed than the other.
 Loose irregular CT is actually fairly common in mammals, but it's quite hard to
see in most H&E preparations because normally it is packed with cells. An
example is the lamina propria of the gut. This is the supporting CT of
the lining of the intestine (slides 40 and 41) upon which the layer of cells
that line the tract are sitting. The cores of villi are made up of this loose
irregular CT, but they are so filled with various types of cells you won't be
able to see the fibers themselves.One example of loose, irregular, collagenous CT is shown at left. This is from the deepest region of the dermis of the skin in slide 35; and really, it's nothing more than the place where the dense irregular CT of the upper dermis beings to thin out. The gradation into the denser region is imperceptible (though not visible in this slide) and maybe this could be characterized as "almost-loose-irregular."
Loose irregular CT is actually fairly common in mammals, but it's quite hard to
see in most H&E preparations because normally it is packed with cells. An
example is the lamina propria of the gut. This is the supporting CT of
the lining of the intestine (slides 40 and 41) upon which the layer of cells
that line the tract are sitting. The cores of villi are made up of this loose
irregular CT, but they are so filled with various types of cells you won't be
able to see the fibers themselves.One example of loose, irregular, collagenous CT is shown at left. This is from the deepest region of the dermis of the skin in slide 35; and really, it's nothing more than the place where the dense irregular CT of the upper dermis beings to thin out. The gradation into the denser region is imperceptible (though not visible in this slide) and maybe this could be characterized as "almost-loose-irregular."
 A better (and more typical) example of loose irregular CT is shown below right. This is the core of an intestinal villus. The collagenous fibers are there, all right, but they're impossible to see against the background of cellular infiltrate. In this location there are typically tens of thousands of cells, all perched on the strands of collagen: mostly plasma cells, but also many macrophages, mast cells, and lymphocytes. There's also a fair amount of smooth muscle in the form on individual strands, anchored to the CT framework. If you scroll over the image the CT area is highlighted.
A better (and more typical) example of loose irregular CT is shown below right. This is the core of an intestinal villus. The collagenous fibers are there, all right, but they're impossible to see against the background of cellular infiltrate. In this location there are typically tens of thousands of cells, all perched on the strands of collagen: mostly plasma cells, but also many macrophages, mast cells, and lymphocytes. There's also a fair amount of smooth muscle in the form on individual strands, anchored to the CT framework. If you scroll over the image the CT area is highlighted.
Regular CT
"Loose regular CT" doesn't exist as a separate type. This makes sense: regular CT's are those in which the fibers run parallel to each other, an arrangement designed to confer tensile strength and stretch resistance. By their nature, any organs that have this sort of structure would be densely packed. the best examples are in tendons and ligaments. Dense regular CT is typified by tendons, the tough collagenous bands that anchor muscles to bones. Slide 46 shows a tendon cut in
longitudinal section, i.e., parallel to the fibers, and slide 31 shows one cut in cross section.
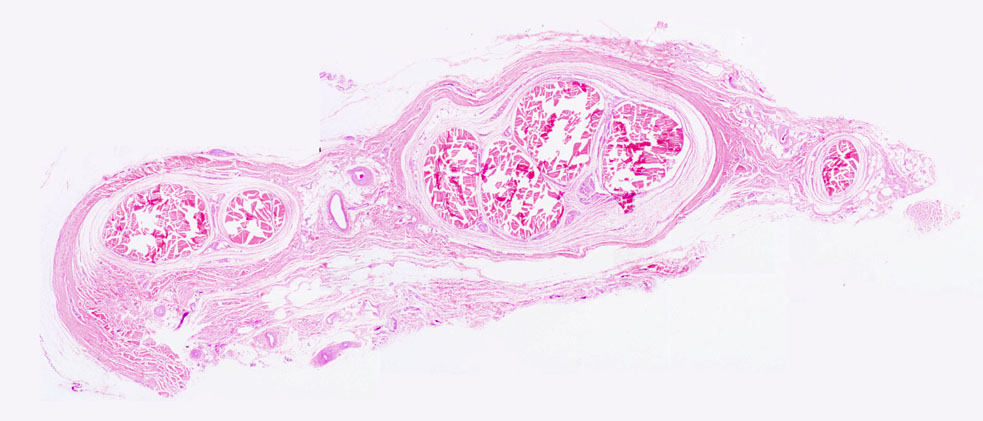
This image is a very low magnification of the tendon on slide 31. The actual tendons are the heavily stained pink areas. These are surrounded by a less intensely stained tendon sheath. The tendon is entirely collagenous in nature, and the fibers in it are laid side by side with their long axes parallel to each other. The fibroblasts/cytes that have made (and are maintaining) the tendon in good condition are sitting in the minute spaces between the fibers, hardly visible as anything more than elongated nuclei. You can readily appreciate
the very regular arrangement of these fibers compared to those of irregular CT.

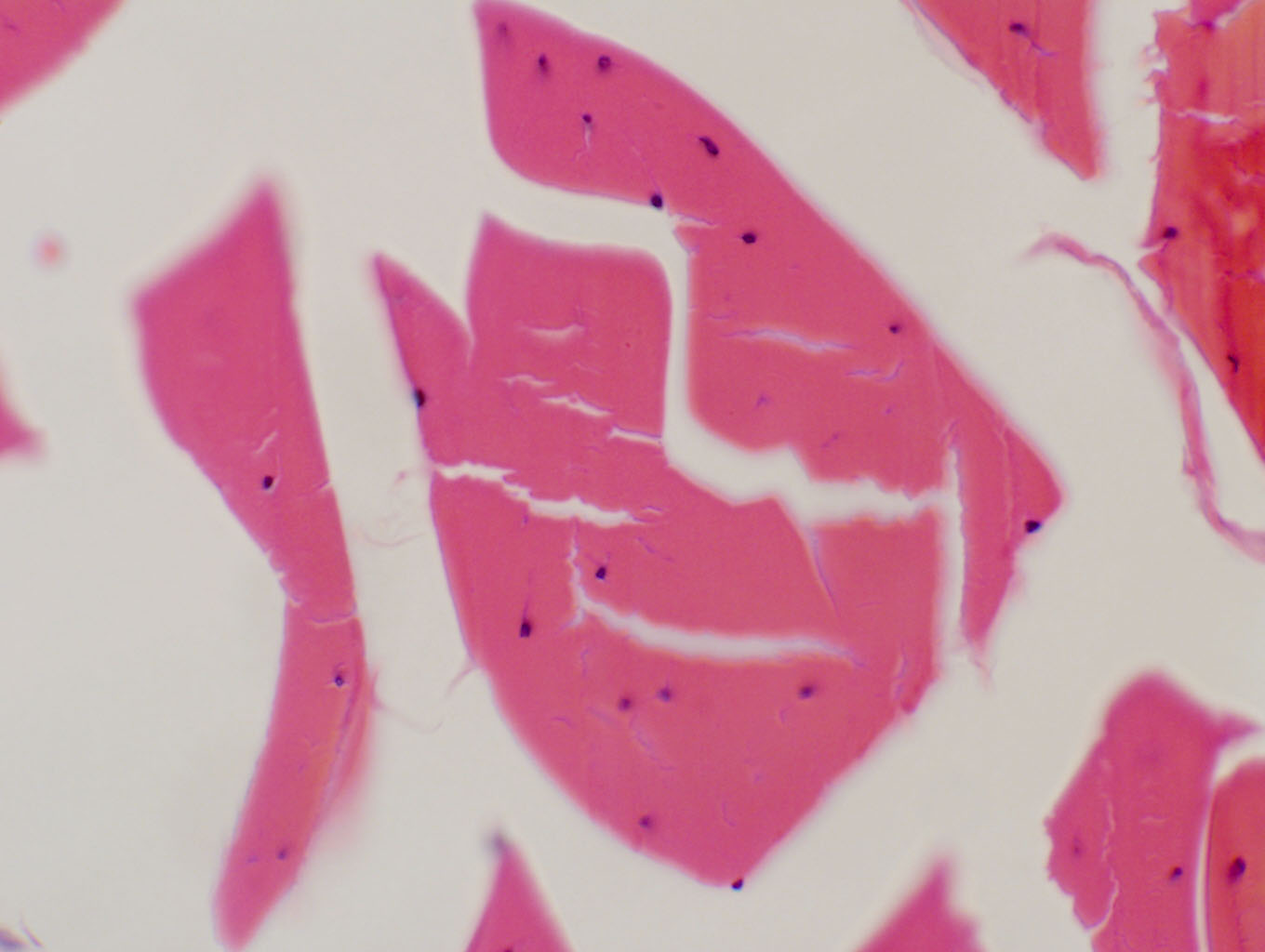 The image at left, from an Achilles tendon on shows these fibers with a "wavy" appearance, which is an artifact. This specimen was fixed after the tendon was cut and so the normal muscle tension was relaxed at the time the tissue was fixed. Still, the parallel course of the fibers is easily visible. The image at right is a high magnification of the cross-sectioned tendon on slide 46. Note that the fibroblast nuclei are between the tightly-packed fibers. It's easier to make out the fibroblasts in both these orientations, but the arrangements of fibers vis-�-vis the cells is more obvious in longitudinal sections.One thing you need to keep in mind, not only for connective tissues but for all others: these are living structures, and they need oxygen and nutrient delivery, as well as waste disposal. There are no "static" tissues in the body: as the old saying goes, "Everybody's gotta have a blood supply," and even seemingly quiescent places like tendon sheaths are no exception.
The image at left, from an Achilles tendon on shows these fibers with a "wavy" appearance, which is an artifact. This specimen was fixed after the tendon was cut and so the normal muscle tension was relaxed at the time the tissue was fixed. Still, the parallel course of the fibers is easily visible. The image at right is a high magnification of the cross-sectioned tendon on slide 46. Note that the fibroblast nuclei are between the tightly-packed fibers. It's easier to make out the fibroblasts in both these orientations, but the arrangements of fibers vis-�-vis the cells is more obvious in longitudinal sections.One thing you need to keep in mind, not only for connective tissues but for all others: these are living structures, and they need oxygen and nutrient delivery, as well as waste disposal. There are no "static" tissues in the body: as the old saying goes, "Everybody's gotta have a blood supply," and even seemingly quiescent places like tendon sheaths are no exception.
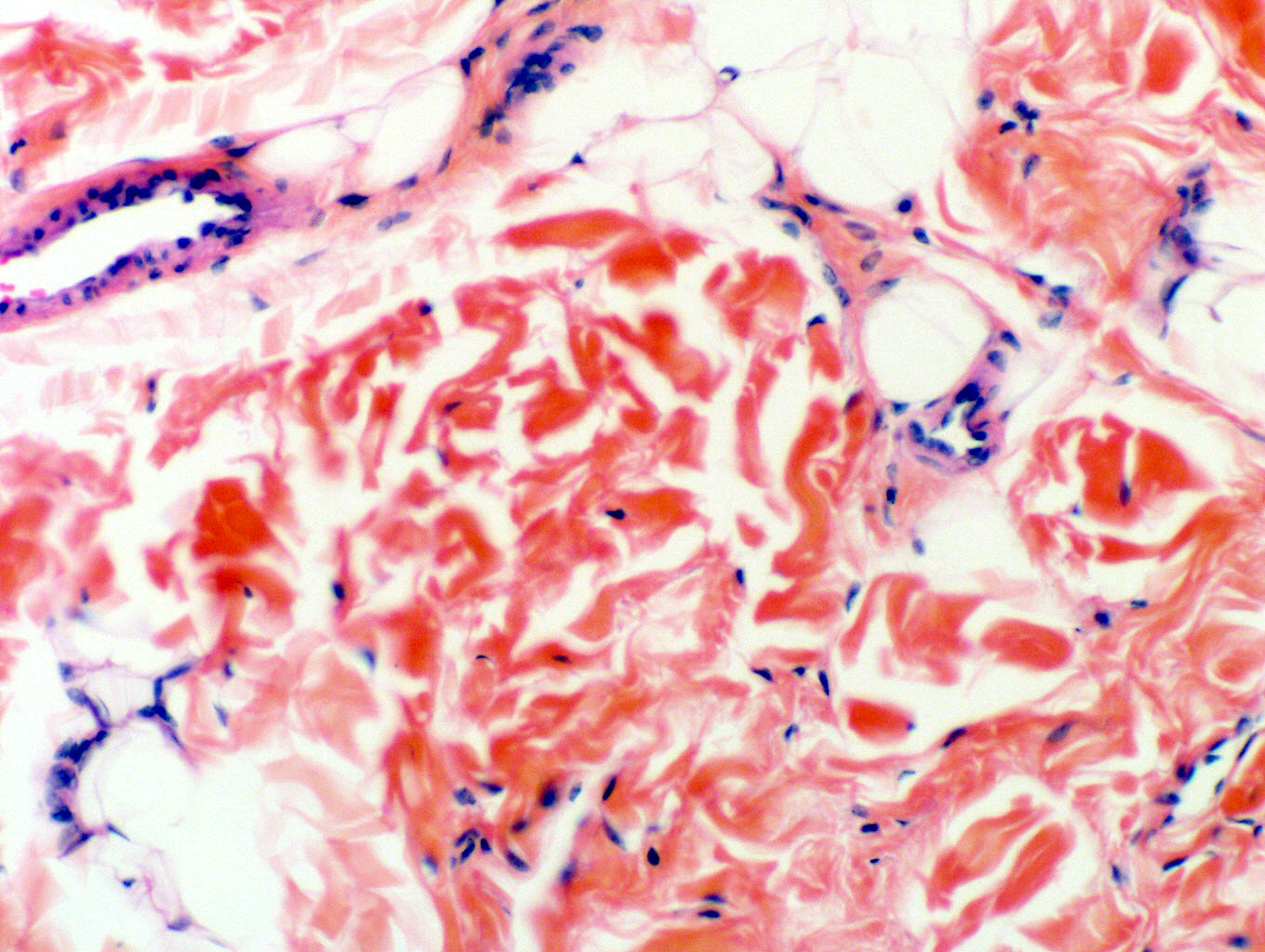 Fibroblasts make non-living fibers, but they're living cells, so you find blood vessels running through CT, even dense CT. The image at left is from a tendon sheath. The dense irregular CT of the sheath is clearly visible, as are the nuclei of the fibroblasts that made it. At right upper center in this field is a small arteriole and a vein accompanying it (veins are large in cross sectional area than their companion arteries, as a rule). Notice also that in this region of mostly collagen fibers, there is nevertheless a fair amount of white adipose tissue visible. White adipose tissue (see below) is commonly found in association with collagenous fibers.
Fibroblasts make non-living fibers, but they're living cells, so you find blood vessels running through CT, even dense CT. The image at left is from a tendon sheath. The dense irregular CT of the sheath is clearly visible, as are the nuclei of the fibroblasts that made it. At right upper center in this field is a small arteriole and a vein accompanying it (veins are large in cross sectional area than their companion arteries, as a rule). Notice also that in this region of mostly collagen fibers, there is nevertheless a fair amount of white adipose tissue visible. White adipose tissue (see below) is commonly found in association with collagenous fibers.
Special CT's
There are several so-called "special" connective tissues, which differ from "proper" CT by
having different proportions of the three components. Some are much more
cellular, and some are mostly matrix. Some are specialized for elasticity, and
some for compression resistance; some are found only in embryos and some only
in some groups of adult animals. All are comparatively limited in distribution
compared to the regular and irregular types discussed above. The special CT's include reticular, elastic, and mucous CT. Also blood, which will be dealt with in Exercise 6.
Reticular CT
Reticular CT has fine reticular fibers in it. These fibers form
very delicate stranded networks, rather than thick bundles, and they provide
the supporting internal stroma for cells in the spleen and other lymphatic
organs. Reticular fibers also invest the outside of small blood vessels and
form part of the basal laminae of epithelial sheets, but fibers in these
locations are difficult to find without special staining. Reticular fibers used
to be considered a separate non-collagenous class, but it's now known that they
are simply a very fine form of collagen, and chemically very similar to other
collagen types. Nevertheless, the classification of reticular CT as a "special" type continues to be used because of its restricted
locations and special role in some organs.
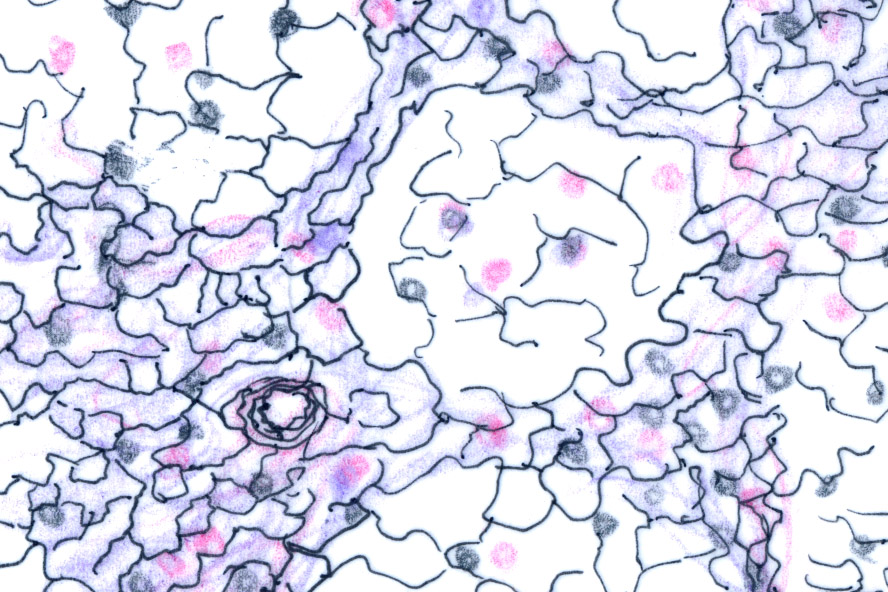
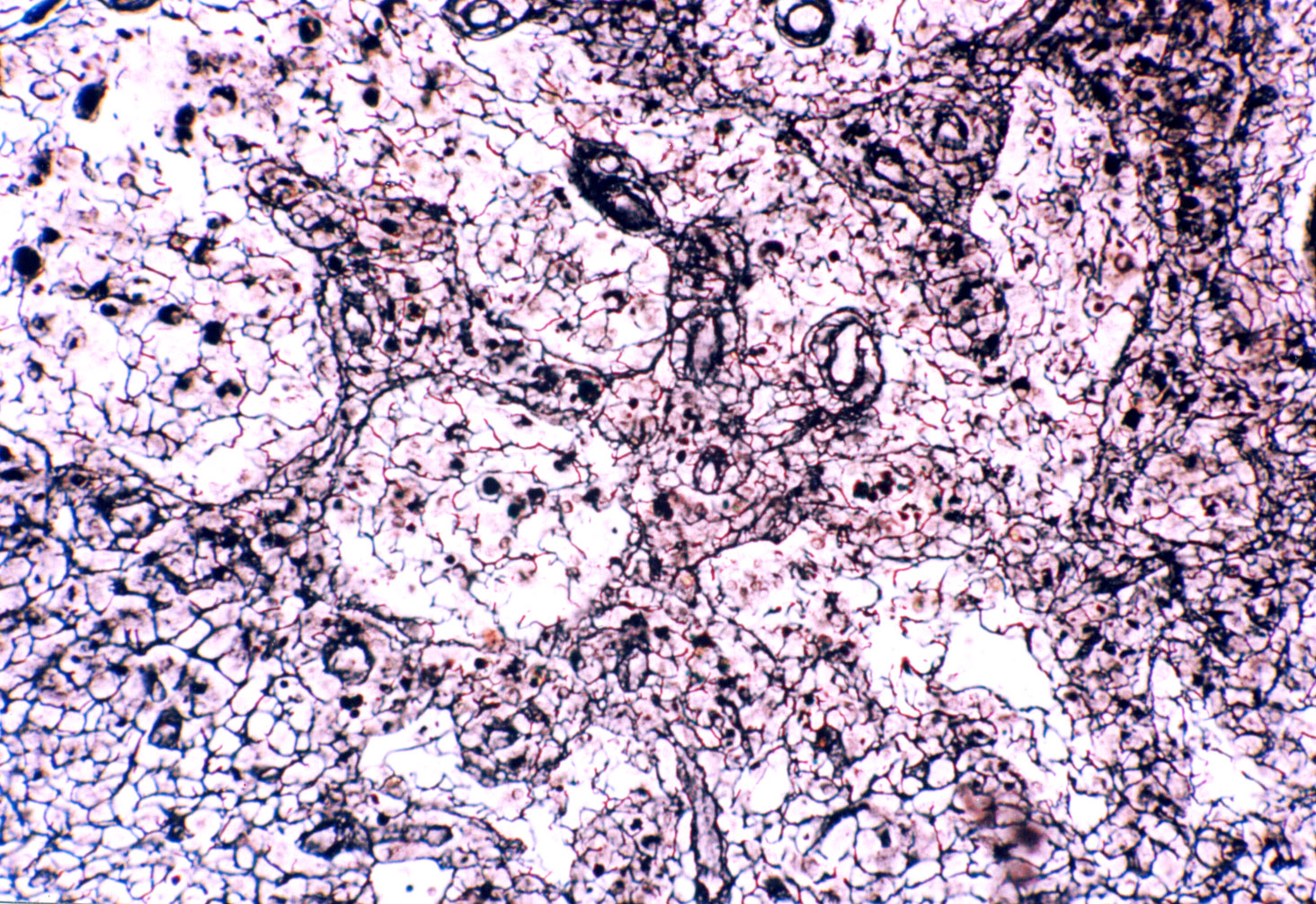
This line drawing and the microscopic image next to it show the nature of the delicate fibers of reticular connective inside a lymph node. The cells normally present are gone and the fibers are very clearly visible. In the field to the right the fibers are seen as thin dark lines. The reticular fiber is really nothing more than a very delicate strand of collagen fiber, less robust than the fibers found in the collagen of skin or in tendons. It's classified as "Type III collagen," and has a few minor differences in the chemical makeup of its strands. This slide is stained with a method that takes advantage of those differences.
 Reticular CT is found in locations where some structural support is needed but the brute strength of normal collagen fibers isn't necessary: for example, the interior volume of organs of the lymphatic system, around small blood vessels, and supporting small nerve processes. The image at right is from slide 29. In the spaces between cells inside
this lymph node fine reticular fibers have been stained with Wilder's reticulum method, a silver impregnation technique more or less specific for reticular fibers. If
you then examine these under higher power, you'll see that they're quite
small and stained black. Some of these fibers are found in the capsule
of the organ as well. The principal role of reticular fibers, here as in other
lymphatic organs, is to provide someplace for cells to sit. Reticular fibers
also serve to form a delicate scaffolding to anchor blood vessels and nerve
fibers to the more robust surrounding CT.
Reticular CT is found in locations where some structural support is needed but the brute strength of normal collagen fibers isn't necessary: for example, the interior volume of organs of the lymphatic system, around small blood vessels, and supporting small nerve processes. The image at right is from slide 29. In the spaces between cells inside
this lymph node fine reticular fibers have been stained with Wilder's reticulum method, a silver impregnation technique more or less specific for reticular fibers. If
you then examine these under higher power, you'll see that they're quite
small and stained black. Some of these fibers are found in the capsule
of the organ as well. The principal role of reticular fibers, here as in other
lymphatic organs, is to provide someplace for cells to sit. Reticular fibers
also serve to form a delicate scaffolding to anchor blood vessels and nerve
fibers to the more robust surrounding CT.
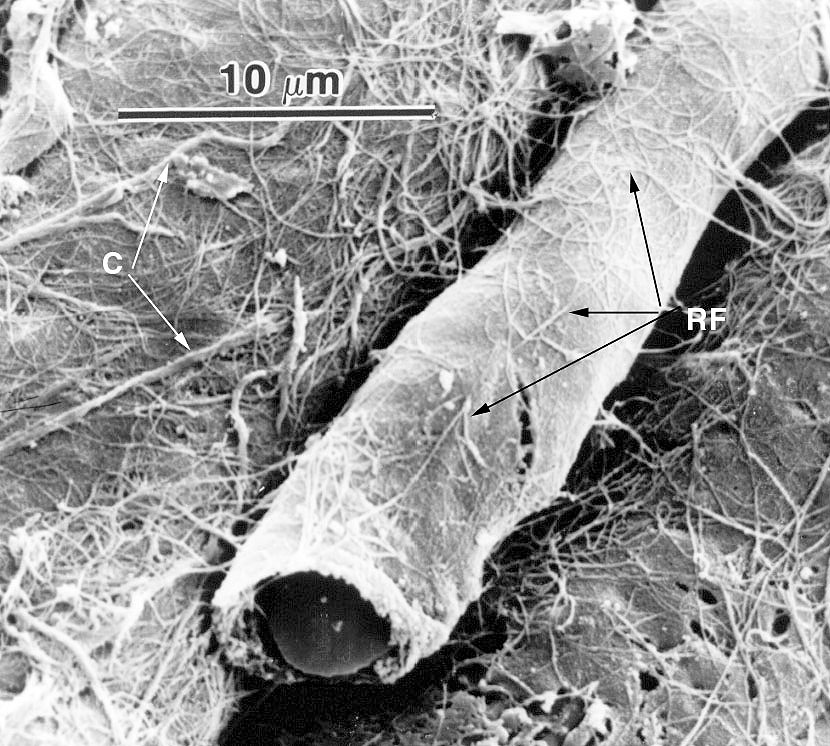
The scanning electron micrograph at left is of a capillary in cardiac muscle, anchored in place by reticular fibers (RF) that connect the outside surface of the vessel to the surround collagen fibers (C) that invest the muscle cells. This "intercellular collagen network" is a common feature of muscle, and indeed of all organs. If this capillary weren't anchored in place it might bend and kink, occluding flow. Anchoring it to the surrounding muscle keeps it patent when the cells around it are contracting and the need for flow is greatest. It's the connective tissue fibers from the reticular level on up to the stoutest tendons that hold everything together and transmit force throughout masses of muscle.
Elastic CT
 Elastic CT, as you might expect, is noted for its springiness. Elastic CT is organized in much the same way as collagenous CT is, with the major difference being the nature of the fibers. It has
some collagen fibers in it, but its fibers are principally made of elastin,
which is chemically different from collagen. Elastic fibers have their own
characteristic amino acid components, desmosine and isodesmosine.
Elastic CT is very limited in distribution and it's not as well developed in
some groups as in others, though all mammals do usually have some of it. The rubbery nuchal ligament is an excellent example of a dense, regular, elastic CT structure. The nuchal ligament (called the supraspinous ligament in gross anatomy texts) helps to raise the head in the horse. When the animal lowers its head to eat or drink, the ligament is stretched. In engineering-speak, like a spring under tension, it's "pre-loaded." Then, when the head is elevated, some of the energy stored in the
Elastic CT, as you might expect, is noted for its springiness. Elastic CT is organized in much the same way as collagenous CT is, with the major difference being the nature of the fibers. It has
some collagen fibers in it, but its fibers are principally made of elastin,
which is chemically different from collagen. Elastic fibers have their own
characteristic amino acid components, desmosine and isodesmosine.
Elastic CT is very limited in distribution and it's not as well developed in
some groups as in others, though all mammals do usually have some of it. The rubbery nuchal ligament is an excellent example of a dense, regular, elastic CT structure. The nuchal ligament (called the supraspinous ligament in gross anatomy texts) helps to raise the head in the horse. When the animal lowers its head to eat or drink, the ligament is stretched. In engineering-speak, like a spring under tension, it's "pre-loaded." Then, when the head is elevated, some of the energy stored in the  ligament is used, lessening the strain on the neck muscles. The ligament is shown above at low magnification: note the very strong similarity to the organization of fibers in the tendon! But while the tendon's function is to resist stretch, this dense, regular elastic connective tissue structure is designed to extend and return to its original length. This slide is stained with orcein, which is specific for elastic CT. These images can be found on slide 32, a nuchal ligament cut in longitudinal section.
ligament is used, lessening the strain on the neck muscles. The ligament is shown above at low magnification: note the very strong similarity to the organization of fibers in the tendon! But while the tendon's function is to resist stretch, this dense, regular elastic connective tissue structure is designed to extend and return to its original length. This slide is stained with orcein, which is specific for elastic CT. These images can be found on slide 32, a nuchal ligament cut in longitudinal section.
At somewhat higher magnification you can see that there's s a reinforcement of collagen fibers. In an orcein stain like this one, they're only faintly visible because they don't react, but they're there. The elastic fibers (EF) provide"springiness" and stretchability, and the high-tensile-strength collagen serves to prevent over-distension. The best mechanical analogy for the nuchal ligament I can imagine is a "bungee cord," those things that are used to hold down tarps and sometimes for jumping off bridges. A bungee cord has a central portion of parallel elastic bands, interwoven with strong thread and overlain by fabric, which prevent it from breaking under tension. This slide is from a horse.Just for the record, while individual strands of elastic fiber are frequently encountered in among the strands of irregular CT's (both regular and irregular) there's no such thing as "loose irregular elastic CT" in mammals. In irregular CT's the predominant fiber type is always some form of collagen, not elastic.
Mucous CT
Mucous CT is generally regarded as an embryonic type, and it's rare or
unknown in adult mammals. However, there are few cases of mucous CT in adult non-mammals: the comb of chickens is filled with it, and so are the wattles of turkeys. (If ever you have the opportunity to examine a turkey's wattles at close range, you certainly should not pass up the chance, it's a once-in-a-lifetime experience.) In this type of CT the matrix is the predominant
element, and fibers are scarce. Cells can usually be seen but they are nowhere
near as numerous as in fibrillar CT's.


Most of the volume of the umbilical cord, seen at left on slide 44, is mucous CT. The image shown here is a low-magnification "orientation" of the cord, cut in cross section. The large pink things in the center are the umbilical arteries and vein, and the stalk of the yolk sac is also present. Surrounding the cord is a delicate epithelial covering, a thin stratified squamous type that's continuous with the surface integument of the fetus.
The apparently empty center of this structure is not: it's filled with mucous CT, and since the umbilical cord is a fetal organ, this can fairly be called "embryonic mesenchyme." The mucous CT
fills the spaces between the large blood vessels. It's mostly ground substance, with few or no fibers; the cells are
elongated and stellate. There's a special name for the
mucous CT of the umbilical cord: it's often called "Wharton's Jelly" in honor of Thomas
Wharton (1614-1673), an English anatomist and physician. The "jelly" in the umbilical cord is derived from mesoderm, so it's angiogenic; in the course of differentiation the mesoderm forms the mucous CT and the vessels through it. These are lost after birth and the termination of the need for placental nutrition.
The scattered dots throughout the field are the fibroblasts which have made all the various components. Matrix material is most glycoprotein, so as you'd expect, this slide would stain very strongly with the PAS reaction. It appears "empty" in an H&E preparation like this one partly because the water soluble "jelly" is mostly lost in processing; and partly because glycoproteins don't stain well with either hematoxylin or eosin. The cells do, however, and have a faintly bluish cast to them, because they're actively secreting material. there are collagen fib ers in here, but they're so few and far between you can't see them at all in the light microscope with any common stain.
Adipose CT
Adipose CT is very cellular and has little if any matrix and few fibers.
As the name implies, its principal function is fat storage. The cells of this
type of special CT are filled with lipid, either in a single droplet or in
small droplets. There are two types, white and brown adipose CT. White
adipose tissue, or "white fat" is very widely distributed,
and quite easy to find.
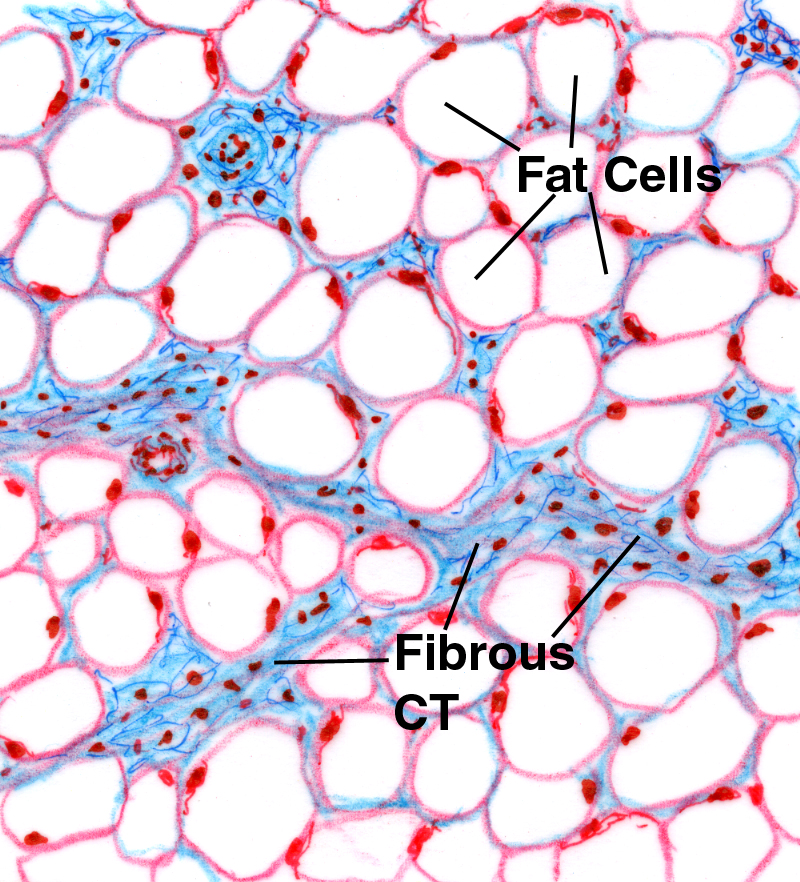

White adipose CT, or "white fat," is quite common. You'll see it as small collections of individual cells, or in larger aggregations, sometimes in globs several kilograms in weight in well-nourished animals. In this low power sketch of a white fat depot (left) and in an actual specimen (right) the typical appearance of this tissue (often referred to as "chicken wire") is pretty much unmistakable. The Mallory's stain used for the sketch has colored the fibrous CT supporting the fat cells blue, and the scanty cytoplasm of the cells is red. The bulk of this type of fat cell is a single large droplet of lipid, which is unstained in such preparations.
White fat serves as a reserve store of energy and as a shock cushion for internal organs. Although it's unfashionable for humans to have a extra pounds of energy reserves in the form of fat cells, in animals it's considered a good thing. A beef carcass is graded in part on the basis of how much fat is present and its distribution. Aggregations of fat cells tend to be distributed throughout collagenous CT's; the fat in the collagen fiber network between muscle bundles is what the beef producer calls "marbling." Well-marbled beef commands a higher price than lean because it's the fat in meat that give it flavor and juiciness.
The total amount of lipid stored in white fat cells fluctuates with diet and condition, but the total number of cells doesn't, because white fat cells are long-lived and normally don't divide. Losing weight by removing fat results from a decrease in the size of the fat droplets in the cells, but not a reduction of cell number. Given a higher input of calories and/or a lower level of energy use, the energy-storage pathway that produces fat is activated again and the cells increase in size.
This is the scientific basis for the "yo-yo diet" phenomenon. People desperate to lose weight have resorted in some cases to surgical removal of fat depots. The theory behind this is that once the cells are gone they don't come back, so there's less tissue volume in which the extra lipid can be stored. In domestic animals, liposuction isn't (yet) a routine procedure, but if people are ever willing to pay for cosmetic surgery on Labrador Retrievers, someone is going to make a killing at it.
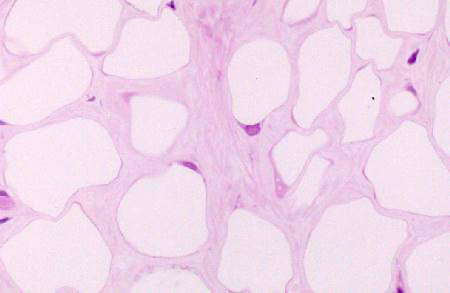 Slide 33 is a chunk of white fat from a pig, and it has the typical "chicken wire" appearance of this tissue. However, you'll probably have better luck with slides 24, 1205 or on of the skin slides (7, 11, 12, or 35). White fat cells occur singly and in small groups in almost all irregular CT areas, especially in the deep layers of the skin. Displacement of the nucleus to one side results in an arrangement that resembles the shape of a high school ring: a larger area with a thin band. At high magnification you should be able to make out this"signet ring" appearance of individual cells. Though the cytoplasm is squeezed into a tiny volume, all the normal organelles are present.
Slide 33 is a chunk of white fat from a pig, and it has the typical "chicken wire" appearance of this tissue. However, you'll probably have better luck with slides 24, 1205 or on of the skin slides (7, 11, 12, or 35). White fat cells occur singly and in small groups in almost all irregular CT areas, especially in the deep layers of the skin. Displacement of the nucleus to one side results in an arrangement that resembles the shape of a high school ring: a larger area with a thin band. At high magnification you should be able to make out this"signet ring" appearance of individual cells. Though the cytoplasm is squeezed into a tiny volume, all the normal organelles are present.
In most H&E slides the actual fat is lost, leached out by the alcohol baths in processing, and only an empty space remains. These cells would be globular in shape if they weren't squeezed up against each other, like soap bubbles clustered into a mass. So the true arrangement is a little different from "chicken wire," but with respect to its two-dimensional appearance in a section that's a pretty good description. White fat has clinical significance. Not only is the level of fat an indicator of general state of nutrition and health, one of the most common tumors of old dogs is the benign lipoma, a tumor of fat. In food animals, quite aside from its importance to the flavor of meat, fat is where pesticide and drug resides usually are stored (most of these compounds are fat soluble and partition preferentially into white fat cells). There's always a "withdrawal period" mandated by law before an animal on certain drugs can be sent to slaughter. Researchers use the "fat burden" of dissolved materials to assess the animals' total load of residues when studying new agricultural or medicinal compounds.
Brown Fat
 Brown fat is considerably more restricted in its distribution than white fat, and unique in its function. It's found in the interscapular region of most mammals at some point in their lives, and it's
especially well-developed in rodents that hibernate, the most famous example being the Eastern Woodchuck, Marmota monax.
Brown fat is considerably more restricted in its distribution than white fat, and unique in its function. It's found in the interscapular region of most mammals at some point in their lives, and it's
especially well-developed in rodents that hibernate, the most famous example being the Eastern Woodchuck, Marmota monax.
Note that in this tissue there isn't
one single large fat droplet inside the cell; rather there are several smaller ones.
This is sometimes called "multilocular fat" as opposed to
"unilocular" white fat. You can find some on slide 113, a cross section of a rat's thorax. Look on slide 113 for a patch of "foamy" looking material in the area between the lungs, dorsal to the esophagus and trachea. Brown fat is brown because it's very heavily vascularized and because its cells contain thousands of mitochondria, which lend an overall brown cast to the fresh specimen. Brown fat is thermogenic tissue: its only function is to generate heat. Energy released from the fat molecules is given off as heat, which is subsequently transferred by the liquid phase of the blood to warm other parts of the body.
Brown fat is exceptionally well developed in rodents, especially hibernating ones like woodchucks and 13-lined ground squirrels. As you might expect, the amount of lipid present (and the size of the 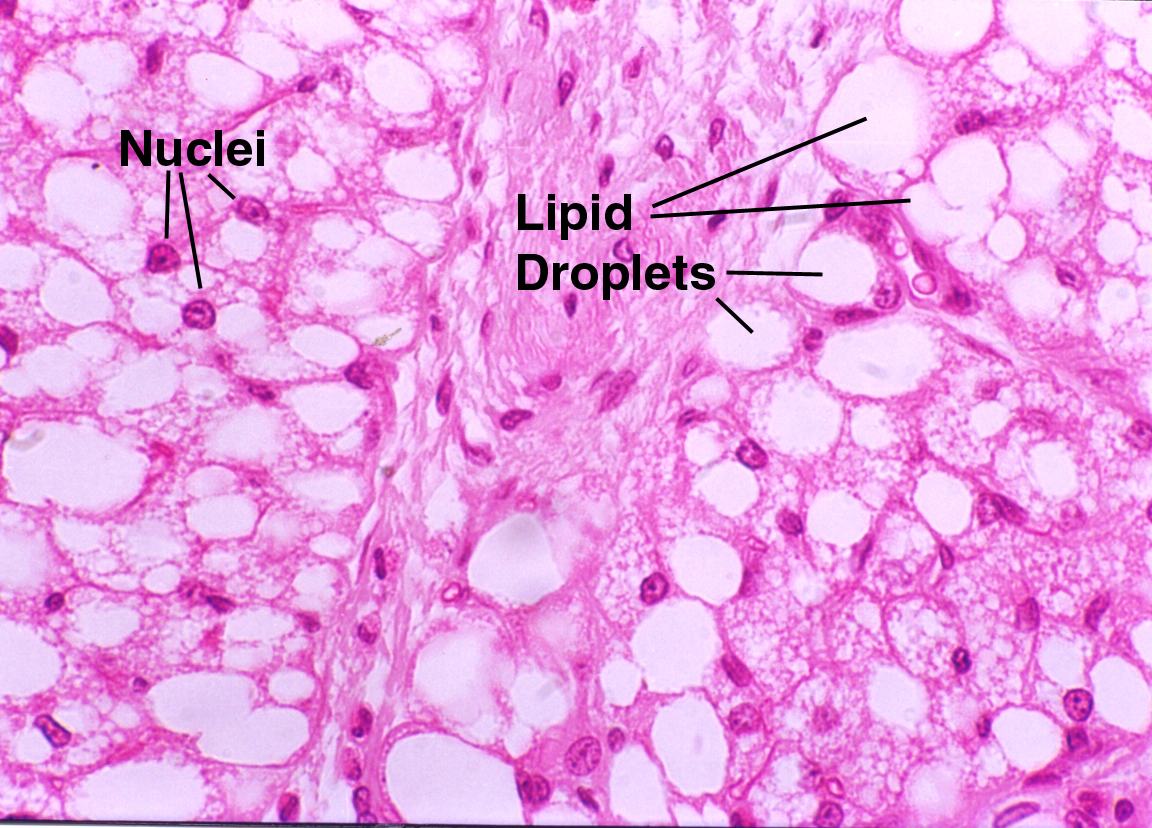
 droplets) varies with nutritional state and in hibernators, the time of year. In the Spring, when woodchucks emerge from their holes, the lipid stores of the brown fat have been used up and one of their physiological requirements is to build them back up again for the next cycle of hibernation and arousal. In non-rodents brown most easily found in young animals where it provides a supplementary source of body heat to organisms without a fully developed thermoregulation capacity.
droplets) varies with nutritional state and in hibernators, the time of year. In the Spring, when woodchucks emerge from their holes, the lipid stores of the brown fat have been used up and one of their physiological requirements is to build them back up again for the next cycle of hibernation and arousal. In non-rodents brown most easily found in young animals where it provides a supplementary source of body heat to organisms without a fully developed thermoregulation capacity.
This sketch of an isolated brown adipocyte schematically depicts the smaller and multiple fat droplets, and the scattered mitochondria, which are actually far more numerous than is represented here. Brown fat is a thermal generator; that is its only function. It has the capacity to decouple the oxidative
phosphorylation of ATP to ADP from the electron transport chain, so that the
energy released by splitting that terminal phosphate group off ATP isn't used
to make other chemical bonds. Instead it's deliberately released as heat into the
surrounding tissue.
Hibernators use brown fat thermogenesis as a way to endogenously raise their
body temperature as they rouse out of their torpor, and so it's best developed
in this group. In non-rodents and non-hibernators, it tends to be present in
young animals (where it serves to stabilize body temperature against
fluctuation to an certain extent) and it diminishes or disappears in later
life, when nervous control of body temperature is fully developed.
Other Cells of CT
We've dealt with fibroblasts, but there are many other cell types in CT, most of
which are common to several CT varieties. Again, these are usually involved in
the supportive and protective functions of this class of basic tissues.
Macrophages

 Macrophages are very common. These phagocytic wandering cells are one of
the major lines of defense against infection, and can be found in most loose CT. You should be able to find macrophages on your own slide 27, in the loose CT of the inflamed salivary gland, from which the image at left comes. Scan the slide at low power looking for brownish pigment. This is hemosiderin and/or lipofuscin, in macrophages that have migrated into the area.
Macrophages are very common. These phagocytic wandering cells are one of
the major lines of defense against infection, and can be found in most loose CT. You should be able to find macrophages on your own slide 27, in the loose CT of the inflamed salivary gland, from which the image at left comes. Scan the slide at low power looking for brownish pigment. This is hemosiderin and/or lipofuscin, in macrophages that have migrated into the area.
These cells contain brown granules of undigested cellular debris and blood cells. Macrophages come in to clean up debris, hemorrhage, and dead cells. Their arrival and activity are normal parts of the sequence of events in the process inflammation and resolution. Macrophages are derived from circulating blood monocytes (see Exercise 6). There is considerable evidence to indicate that the monocyte isn't a "blood cell" at all: it just uses the blood as a transport mechanism to get where it has to go, then leaves the vascular system for the connective tissue space, where it differentiates into a macrophage. 
Macrophages are vitally important components of the immune system's mechanism, conferring the capacity to deal with particulate contaminants. Some organs have a certain number of "resident" macrophages, present all the time, to deal with routine contaminants. This is true of almost all lymphatic organs. The example at left is a large and active resident macrophage in a lymph node. (Other, smaller, less active macrophages will be indicated when you scroll over this image.) Macrophages are very "pleiomorphic," i.e., their appearance varies with physiological state and activity; so be prepared to see them without the large amounts of ingesta visible here and in the previous example. The lung in mammals is an especially good example of organ-resident macrophages. Millions of macrophages crawl about on the lung surface with an amoeboid motion, gobbling up bits of debris and bacteria that are inhaled, to keep the surface of the lung essentially sterile. Resident macrophages can also be found in the liver, the spleen, and the lamina propria of the digestive tract. When needed to deal with a specific problem, more macrophages can be produced. Precursor cells in the bone marrow differentiate as monocytes which are "recruited." They leave the marrow, enter the blood and migrate to the site of infection as monocytes, immediately differentiate, and get to work. You should be able to see these easily on slide 111; also see Exercise 25.
Plasma Cells
Plasma cells are a very common and very important cellular component of
CT. They're also present in inflammatory sites and in lymphatic organs. They can be found most easily in the region immediately underlying the
absorptive epithelium of the gut, which you'll see on slide 40.
The plasma cell is the most important of the "wandering" cell types of CT of the gut: they're what was once a "lymphocyte" (see Exercise 6). The plasma cell is a tissue resident form of one
of the cells of the circulating blood, the lymphocyte. Its function in tissue
is the production of antibodies. Lymphocytes becomes "transformed" into a plasma cell by meeting its specific antigen; this encounter triggers a process of RER proliferation and active antibody production. Transformation normally occurs in the interstitial spaces of CT. It's especially likely to occur in the loose collagenous CT that supports the epithelium of the gut, because the epithelial barrier there is fragile and exposure to antigens is a normal occurrence. That's why plasma cells are so easy to find in the loose irregular CT of the gut; but they can also be seen in many other locations where antigens are present. 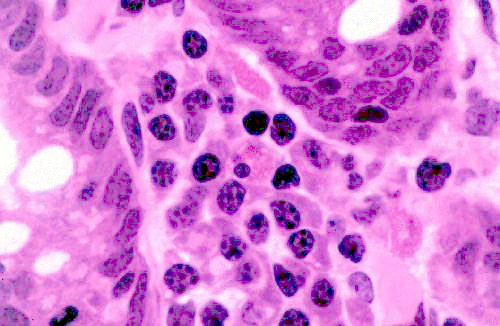
The plasma cell is easily recognized by its
chunky, squarish shape, intensely stained cytoplasm (which is quite basophilic,
since these cells are making antibodies at a terrific rate) and by its characteristic "clock face" pattern of chromatin
distribution in the nucleus. Since antibodies are proteins, this cell in
its "active" state shows all of the features expected of any protein
secreting cell. These cells are packed solid with RER, accounting for their
deep basophilia. As you'd expect from a protein-making cell, they have a good deal of basophilia.
Another classic hallmark of the plasma cell is the so-called "clock face" pattern of nuclear chromatin. The prominent nucleolus in the center is surrounded by clumps of heterochromatin. The nucleus is usually eccentrically located in these squarish, chunky cells. (There's a mast cell in this field, too. Its nucleus is much smaller and denser than those of the plasma cells). Keep in mind that the transitions from lymphocyte to plasma cell and back to a lymphocyte are continuous; that is, there are intermediate stages all along the path in both directions, and you may see not-quite-plasma cells and not-quite lymphocytes as well as the definitive beginning and end stages of the process.
Mast Cells


Mast cells are very difficult to see in wax impregnated H&E
preparations. The sketch at left shows the basic characteristics of this cell type. There's a single nucleus, and numerous large and obvious granules in the cytoplasm. These characteristic granules are easily lost in conventional processing of wax sections. In plastic embedded materials, however, you can find them
easily, because the embedment isn't removed in preparation and the granules are therefore preserved and stained.
The water soluble granules are invariably lost in preparations of wax embedded material because they're washed away when the specimen is hydrated for staining. In plastic sections the embedding material is permeable to water (which wax isn't) and so the granules aren't lost. Mast cells are very widely distributed in most CT's. The mast
cell's functions are not completely clear, but they are known to be involved in
allergic reactions. Mast cells have coarse intracellular granules, their best
identifying characteristic in slides. The granules contain heparin and histamine (and in some species also serotonin).
Mast cells "degranulate" or release their contents into the intercellular space
as part of an allergic reaction and sometimes in response to neural
stimulation.You should find them very easily on slide 40, from which the image at right was made. Look for an oval cell with a small nucleus and granulated cytoplasm.
They'll stand out pretty nicely against the eosinophilic background of the CT, and you'll also find them in the cores of villi, in the same location as plasma cells. The fact is, mast cells can be found anywhere in almost any CT.They somewhat resemble eosinophils (which are also found here, see Exercise 6) but the nuclear shape
gives them away. the eosinophil nucleus is classically bi-lobed by the nucleus of the mast cell is not.
The mast cell is the tissue-resident form of the basophil, a cell type found in circulating blood. This is based
on some hard data regarding the contents of the granules in both cell types,
but the theory has yet to be definitively proven. It's important to point out that there's actually a pretty specific stain for mast cells, and that these cells are sometimes involved in tumor formation. Below and left you'll see an image of mast cells, from a mast cell tumor. This slide has been stained with toluidine blue, which is pretty specific for mast cells. The granules are very obvious, and it's pretty easy to distinguish the mast cells in a preparation like this.


As a comparison of the two stains, I've put an image of mast cells revealed with TB and H&E side by side. Don't be fooled by the eosinophilia of these cells into thinking they're eosinophils! The giveaway is the single nucleus and the coarse, very obvious granules.
Home Page
Lab Exercise List