VM8054 Veterinary Histology
Exercise 27: Male Reproductive System
Author: Dr. Thomas Caceci
He that is wounded in the stones, or hath his privy member cut off,
shall not enter into the congregation of the Lord.
Deuteronomy XXIII:1
 Objectives for this Exercise
Objectives for this Exercise
PRE-RECORDED LECTURE FOR THIS EXERCISE
Male Reproductive Organs
The male reproductive system consists of the testes and their associated excurrent ducts to conduct sperm to the outside. Usually also included in this system are various male accessory glands which produce non-sperm components of the ejaculate; and that portion of the
urethra which is used for the transport of semen. It also includes the
intromittent organ, the penis.
The testes are both "exocrine" and endocrine in nature. They produce
both a cellular "secretory product" in the form of sperm, and the
hormone testosterone as a true endocrine secretion. I think it's stretching the definition of an exocrine gland to classify the testis as one, you'll sometimes see it referred to in textbooks as a "cytocrine" (i.e., cell-secreting) exocrine organ, so it's worth mentioning. There's no question about it's endocrine function, however. These two products
come from different regions of the testes, which we'll examine separately.
The testes of most mammals are located extra-abdominally in the scrotum. This is
actually a temperature controlling device. In most mammals spermatogenesis
proceeds evenly and smoothly at an optimal temperature slightly below the
core body level. The role of the scrotum is to maintain that temperature. When
the ambient temperature drops, the testes are
pulled up by muscular action towards the warmth of the body cavity, and when it
rises, the muscle relax, allowing the testes to descend and remain cool. Almost
all terrestrial mammals have extra-abdominal testes.
Testis
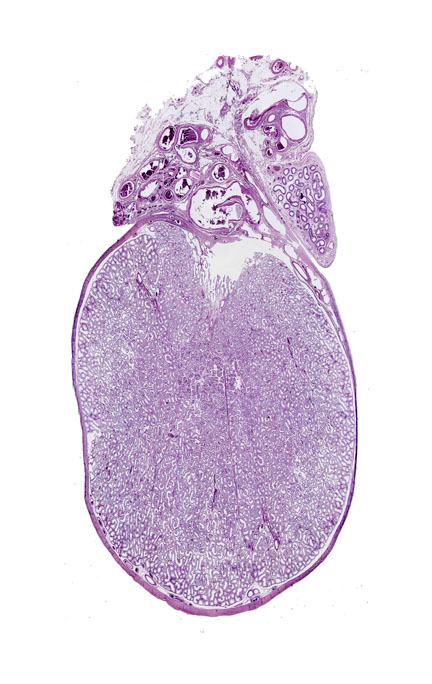 Start with slide 51. The outer border of the testis is demarcated by a thick
band of dense collagenous connective tissue, the tunica albuginea. The
tunica albuginea has a good many blood vessels running through it, including
some rather large arteries. This is to be expected, since the inner portions of
the testis require good circulation, and the blood supply has to come through
the outer tunic. From the tunica albuginea, small septa subdivide the
cavity of the testis into smaller compartments. Septation is usually incomplete
in domestic animals.
Start with slide 51. The outer border of the testis is demarcated by a thick
band of dense collagenous connective tissue, the tunica albuginea. The
tunica albuginea has a good many blood vessels running through it, including
some rather large arteries. This is to be expected, since the inner portions of
the testis require good circulation, and the blood supply has to come through
the outer tunic. From the tunica albuginea, small septa subdivide the
cavity of the testis into smaller compartments. Septation is usually incomplete
in domestic animals.
 The section of testis at left is a cut through the approximate midline; at right a higher magnification image shows a little more detail of the peripheral regions.
The section of testis at left is a cut through the approximate midline; at right a higher magnification image shows a little more detail of the peripheral regions.
In addition to the testis proper the left image includes a piece of the spermatic cord and part of the epididymis, both extra-testicular structures. It shows quite well the very thick tunica albuginea of the organ. This "white coat" (so named from its appearance in fresh gross anatomical specimens) is a dense connective tissue envelope. From it emerge septae that subdivide the internal volume of the testis. The degree of internal septation is species variable, and in most domestic animals there isn't much of it. Here you can see a few small incursions of CT off the tunica albuginea. The bulk of the volume of the testis is filled with the seminiferous tubules: There may be several hundred to a thousand or so per testis. Each one consists of a long loop whose outlet is via channels in the tunica albuginea. We'll examine the contents of the seminiferous tubules below.
Seminiferous Tubules
 The bulk of the testicular tissue is the seminiferous tubules which are present in astonishing quantity. A human testis may have 800-1600 tubules, with an aggregated length of about 600 meters: that's a shade over 1950 feet, which is a good deal longer than the Empire State Building is high. The figure is much higher for
large animals like boars and bulls. Obviously this is a system designed for massive production of sperm on a constant basis.
The bulk of the testicular tissue is the seminiferous tubules which are present in astonishing quantity. A human testis may have 800-1600 tubules, with an aggregated length of about 600 meters: that's a shade over 1950 feet, which is a good deal longer than the Empire State Building is high. The figure is much higher for
large animals like boars and bulls. Obviously this is a system designed for massive production of sperm on a constant basis.
The outer wall of each seminiferous tubule
is a single layer of boundary tissue cells similar in some ways to
smooth muscle. They're sometimes called myoid cells or peritubular
contractile cells, and in at least in some species (e.g., rodents)
they are known to be contractile, though contractility hasn't been proven of all animals. These "myoid cells" have some characteristics of smooth muscle, but structurally they're flattened into a shape reminiscent of squamous epithelium, and butted edge to edge to form the wall of the tube. The contractions of the boundary cells appear to be spontaneous: nervous
stimulation isn't required. Between the seminiferous tubules—in the interstitial region—you
will also find blood vessels of the peritubular capillary network, and the interstitial
cells, discussed below.
Cell Types in the Seminiferous Tubules: The Sertoli or Sustentacular Cell
The lining cells of the tubules comprise the seminiferous epithelium. Some of them are the source of sperm and some are
there to sustain the production.
The Sertoli cell or sustentacular cell is easily identifiable in H&E preparations by its
nucleus. This cell type is not part of the sperm cell
line. Instead it's a "nurse" cell that sustains the proper environment
for spermatozoa to develop. The Sertoli cells sits on the outermost portion of the tubule, right up against
the boundary layer: Its nucleus will be slightly separated from the tube border, pale-staining, with an oval to pyramidal shape, and one or two
prominent nucleoli. Although these cells have a very extensive and branching
cytoplasmic structure, little of the Sertoli cell's true extent can be seen in
the light microscope. The extensive branching nature of the cell's cytoplasm
and its role in isolating the haploid components of the system from the rest of
the body were not understood before the advent of the EM.
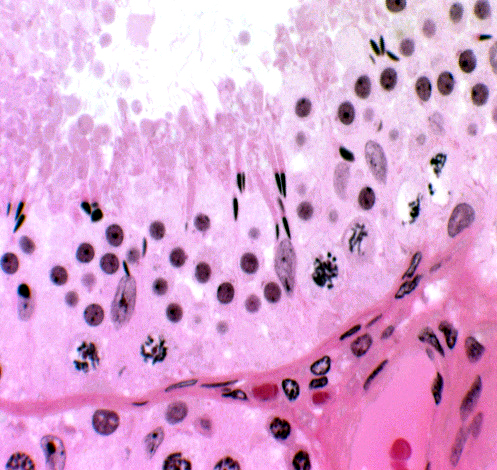 This field shows several Sertoli cells, and also demonstrates their relationship to the wall of the tubule and to other cells in the seminiferous epithelium. Sertoli cells are really the only true "resident cells" of the seminiferous tubules. The various stages of spermatozoal development all are destined to mature and be used up, but Sertoli cell is present before puberty and remains in place until death.
This field shows several Sertoli cells, and also demonstrates their relationship to the wall of the tubule and to other cells in the seminiferous epithelium. Sertoli cells are really the only true "resident cells" of the seminiferous tubules. The various stages of spermatozoal development all are destined to mature and be used up, but Sertoli cell is present before puberty and remains in place until death.
They're a permanent population that doesn't normally divide and proliferate, unlike the germ line around them. (They can occasionally become neoplastic, producing a Sertoli cell tumor.) In this image the distinctive appearance of the Sertoli cell's nucleus, with its rhomboid inclusion and vesicular nucleoplasm, is clearly evident.
The cytoplasm isn't visible without special stains, but using these reveals that the Sertoli cell's overall shape is somewhat like a tree. The early stage cells in the line of sperm development are located near the base of the "trunk" and as they develop into later stages, the spermatocytes are moved up into the "branches" of the tree. By the time they get to the part close to the tubule lumen, they have completed their meiotic division and become spermatids, haploid cells undergoing the final morphological transformation into sperm.
A major role of the Sertoli cell is to define two separate and physiologically different compartments in the seminiferous tubule. The basal compartment, near the periphery, is sealed off from the luminal compartment by processes of adjacent Sertoli cells that reach out and make contact: where the processes are in contact they form occluding junctions. Below the junction, in the basal compartment, are located the diploid spermatogonia. Once these commence division, the cells send out new processes to "undermine" the forming spermatocytes, and seal off a new layer below them, then break the barrier above. This effectively transfers the haploid cells to the luminal side and prevents the diploid ones from being exposed to it.
 Why is this elaborate mechanism for separating the tubule into two compartments necessary? The basal compartment has access to the blood stream and all the immune system components it carries. The haploid gametes are similar to the diploid cells but different enough to be "non-self." There's a risk that the immune system might attack them, and produce unwanted systemic autoimmune reactions as a complication.
Why is this elaborate mechanism for separating the tubule into two compartments necessary? The basal compartment has access to the blood stream and all the immune system components it carries. The haploid gametes are similar to the diploid cells but different enough to be "non-self." There's a risk that the immune system might attack them, and produce unwanted systemic autoimmune reactions as a complication.
Another very good reason is that the haploid gametes are physiologically "compromised" and the sperm especially lack the capacity to deal with harmful materials that might be borne in the blood. So for everyone's protection, the blood-testis barrier is created by the occluding junctions of adjacent Sertoli cells. Maintenance of two separate
compartments allows the different stages of the sperm cycle can exist under
optimal conditions.
Cell Types in the Seminiferous Tubules: Stages of Sperm Development
The seminiferous epithelium contains numerous stages in the developmental sequence that leads to mature spermatozoa. The earliest of these is the spermatogonium, the stem cell from which the rest of the cell types are derived. Some of the spermatogonia remain
undifferentiated, to ensure a continuing supply of stem cells. Others undergo meiosis, become haploid spermatocytes, which then complete morphological differentiation into spermatozoa.
 Spermatogonia are basally located (i.e., at
the periphery of the tubule) and can be identified by their densely stained,
round nuclei. These are diploid cells, and they're in the basal compartment created by the Sertoli cells.
Spermatogonia are basally located (i.e., at
the periphery of the tubule) and can be identified by their densely stained,
round nuclei. These are diploid cells, and they're in the basal compartment created by the Sertoli cells.
Primary spermatocytes are the initial product of spermatogonial
maturation; you will see these as large cells with a round nucleus filled with
clumped chromatin material. Many of them should be seen in various stages of
the meiotic cycle.
Although secondary spermatocytes are undoubtedly
present, these are very short-lived and you're unlikely to tell it from a primary spermatocyte. By the time cells have progressed to the spermatocyte stage meiosis is complete; hence secondary spermatocytes are haploid and located in the luminal compartment.
 The secondary spermatocytes further transform themselves into spermatids. Spermatids are well on the road to final morphological differentiation. They're located
much closer to the lumen of the tube, resting in the branches of the Sertoli cell "tree." Spermatids will be seen in different stages of maturation. Some will be chunky and squarish-looking cells with nuclei beginning to get
quite dense, as at left. Others more advanced will have tails beginning to
develop. You may also see stages in which the excess cytoplasm is being cast
off. A "tailed spermatid" is a fairly common sight.
The secondary spermatocytes further transform themselves into spermatids. Spermatids are well on the road to final morphological differentiation. They're located
much closer to the lumen of the tube, resting in the branches of the Sertoli cell "tree." Spermatids will be seen in different stages of maturation. Some will be chunky and squarish-looking cells with nuclei beginning to get
quite dense, as at left. Others more advanced will have tails beginning to
develop. You may also see stages in which the excess cytoplasm is being cast
off. A "tailed spermatid" is a fairly common sight.
 The spermatids at right have virtually completed their spermiogenesis, are almost ready to be cast off into the lumen to begin their journey. A veritable forest of tails waves above them, and the accumulated cast off cytoplasmic droplets are below. Beneath the maturing sperm, another wave of spermatids is beginning the process of differentiation.
The spermatids at right have virtually completed their spermiogenesis, are almost ready to be cast off into the lumen to begin their journey. A veritable forest of tails waves above them, and the accumulated cast off cytoplasmic droplets are below. Beneath the maturing sperm, another wave of spermatids is beginning the process of differentiation.
Spermatogenesis proceeds in different stages at different parts of each tubule, so that there are always some at each stage of development. Spermatogenesis is a continuous process, unlike the cyclic process of oogenesis. During the breeding season, male mammals are always supplied with matured sperm, and in most cases will breed with any willing female. The process of continuous spermatogenesis assures that such breedings aren't merely "going through the motions," and that there's at least some chance a pregnancy will result.
Interstitial Cells
 In the regions between the seminiferous tubules (i.e.,
outside the adjacent boundary layers) run small amounts of CT, and in there also will be found many capillaries, as is true of all such CT regions. Another population of cells is located here, one not part of the germ line. These small groups are cell which produce the male
steroid hormone testosterone. Their location gives them their name: interstitial cells (or Leydig
cells for Franz von Leydig, 1821-1908, at left; a German anatomist). They're
cuboidal in shape and present in groups of 2-20 cells. You should be able to
make these cells out without difficulty on slide 51. e
In the regions between the seminiferous tubules (i.e.,
outside the adjacent boundary layers) run small amounts of CT, and in there also will be found many capillaries, as is true of all such CT regions. Another population of cells is located here, one not part of the germ line. These small groups are cell which produce the male
steroid hormone testosterone. Their location gives them their name: interstitial cells (or Leydig
cells for Franz von Leydig, 1821-1908, at left; a German anatomist). They're
cuboidal in shape and present in groups of 2-20 cells. You should be able to
make these cells out without difficulty on slide 51. e
 Interstitial cells, like any hormone-secreting cells, have to have close association with blood vessels, so placing them in the spaces between seminiferous tubules makes sense. Furthermore, their secretion is necessary for normal testicular development and for the processes of spermatogenesis and spermiogenesis to take place. Local diffusion serves the needs of the cells in the seminiferous tubules, and blood borne distribution of testosterone triggers the development of male secondary sex characteristics such as growth of facial hair and changes of body morphology. The pituitary hormone ICSH (Interstitial Cell Stimulating Hormone) is the trigger for the interstitial cells to start making testosterone at the onset of puberty. Testosterone is a steroid hormone, so these cells have a good deal of smooth endoplasmic reticulum, when observed in the electron microscope. Their nuclei have the vesicular staining pattern and the prominent nucleolus typical of other secretory cell types.
Interstitial cells, like any hormone-secreting cells, have to have close association with blood vessels, so placing them in the spaces between seminiferous tubules makes sense. Furthermore, their secretion is necessary for normal testicular development and for the processes of spermatogenesis and spermiogenesis to take place. Local diffusion serves the needs of the cells in the seminiferous tubules, and blood borne distribution of testosterone triggers the development of male secondary sex characteristics such as growth of facial hair and changes of body morphology. The pituitary hormone ICSH (Interstitial Cell Stimulating Hormone) is the trigger for the interstitial cells to start making testosterone at the onset of puberty. Testosterone is a steroid hormone, so these cells have a good deal of smooth endoplasmic reticulum, when observed in the electron microscope. Their nuclei have the vesicular staining pattern and the prominent nucleolus typical of other secretory cell types.
Excurrent Ducts: Rete Testis, Epididymis, and Ductus Deferens
Sperm, once cast off into the lumen of a seminiferous tubule, can't just stay there: they have to be up and at 'em, engaged in the Great Quest, which means they have to leave. They do this through a series of channels that run through the CT of the testis' wall, thence to some short convoluted tubules to a single long one that leads eventually to the urethra.

The tunica albuginea has a thickened region in its centerline, the mediastinum testis. Through this run tortuous channels lined with a low cuboidal epithelium, rather like channels in a river delta. This is the rete testis. The seminiferous tubules discharge their spermatozoa into these channels, which conduct them to the 7-15 efferent ducts. The efferent ducts are larger than the channels through the mediastinum, and collectively lead into a long, extremely coiled single duct, the ductus epididymidis. This duct fills the extra-testicular organ, the epididymis. These areas will be highlighted above in different colors if you scroll over the image; it will help to keep you oriented in the discussion below if you keep these three segments of the excurrent path in mind.
 Rete Testis
Rete Testis
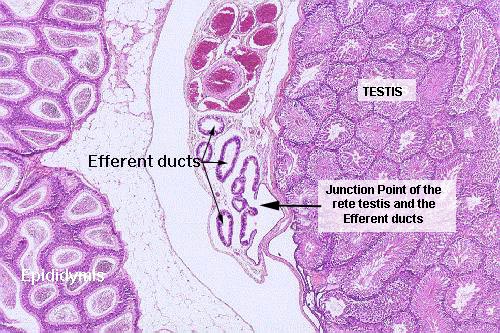
Slide 241 has on it a testis from a hamster. The epididymis is also present as two
crescentic areas on each side of the testis. The connection between the rete testis and the efferent ducts can be made out on this slide. The exit from each seminiferous tubule is "guarded" by Sertoli cells, where the ends of the tubules join the rete.
Between the epididymis and the testis on slide 241 there's a clear area: a set of small ducts and an
artery surrounded by veins can be seen, and at about
10x the portion of the duct system closest
to the testis is seen to be lined by simple cuboidal epithelium. This is the rete, where it emerges from the mediastinum to lead into the adjacent efferent ducts. The epididymis is to the left in this field, and part of the spermatic cord (with large numbers of blood vessels) is at the top center. The junction of the outflow from testis into the efferent ducts is labeled.
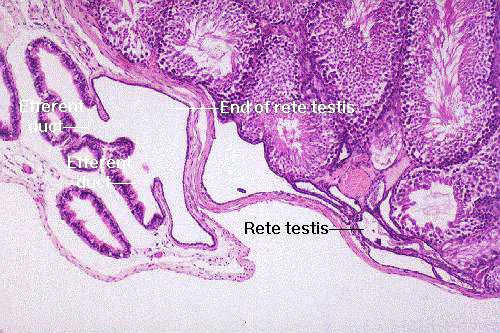 The relationship of these two is better seen in this higher magnification image. There's a portion of the
intra-testicular rete testis in the lower right portion of this field. The retial tubules coalesce into a sort of sinus, into which sperm move, and from which they depart via the efferent ducts. Notice that there's a very abrupt change in the nature of the epithelial lining from the low cuboidal of the rete to the rather elaborate lining of the efferent ducts at the junction.
The relationship of these two is better seen in this higher magnification image. There's a portion of the
intra-testicular rete testis in the lower right portion of this field. The retial tubules coalesce into a sort of sinus, into which sperm move, and from which they depart via the efferent ducts. Notice that there's a very abrupt change in the nature of the epithelial lining from the low cuboidal of the rete to the rather elaborate lining of the efferent ducts at the junction.
Efferent Ducts
The rete testis opens into seven or eight efferent ducts. These relatively short and highly coiled ducts are lined by a pseudostratified columnar epithelium. Many (but not all) of the epithelial cells lining the duct,
especially the taller ones, are ciliated. These are true cilia capable of movement.
The cilia help propel the spermatozoa
along their journey. This is needed because even though they're morphologically normal, the sperm's capacity for independent movement hasn't yet developed.
 The efferent ducts are derived from pre-existing embryonic structures. The drainage ducts of the mesonephros, the so-called "middle kidney" develop into these structures in the term fetus.
The efferent ducts are derived from pre-existing embryonic structures. The drainage ducts of the mesonephros, the so-called "middle kidney" develop into these structures in the term fetus.
Not all of the mesonephric ducts get used, however: some are never connected to the developing gonads. For the most part the unused ones degenerate with the rest of the mesonephros, but alongside the "successful" ones that metamorphosed into efferent ducts, you may see a few of these aberrant ducts. They lead nowhere, connect nothing, and have no significance: they're just vestiges of a long-vanished mesopnephros without functional significance.
Epididymis


Now let's go back and examine the epididymis itself. The "tubules" in
it are really one very long, monstrously coiled tubule (up to 200 yards
long in some species). Although in sections it appears to be a large number of separate tubes, it isn't: it's one tube, cut through at numerous places. This can easily be shown by dissection of gross specimens. It receives input of sperm from the efferent ducts and it acts as a storage and maturation facility for them. Usually the lumen of this tube is filled with sperm that
have been released from the seminiferous tubules "upstream."
The epididymis is lined by a columnar epithelium adorned at its surface with large microvilli. Unfortunately, the true nature of these structures wasn't known before the invention of the electron microscope. They can easily be seen in a light microscope, where they resemble the cilia found in other parts of the body. This similarity in appearance led to them being dubbed "stereocilia," a name that's still used today even though they are not cilia in any sense of the word. Cilia have internal microtubules and are capable of movement: "stereocilia" do not have the internal structure and they can't move at all. They're filled with cytoplasm, and they're a structure for absorption, not movement of fluids.

The epididymis is an absorptive and secretory organ: its cells take up excess fluid that's passed out of the efferent ducts, hence the need for greater surface area.
Sperm have to spend some time in the epididymis to be "capacitated" before they become capable of fertilizing an oocyte. It's the lining epithelium that does this. It produces glycoprotein material that coats the sperm. The secretory nature of the epithelium is revealed by the very large and easily-seen Golgi apparatus profiles in the cells, located between the free surface and the nucleus. If an animal engages in repeated copulations with only very short intervals between, not only does sperm count in the ejaculate decline (because the upstream supply is being depleted), but the fertilization potential is lowered because more immature sperm are included.
 The image on the right is from the initial upstream" end of the epididymis, the head or caput portion. The epithelium here is of "festooned" appearance, i.e., of varying height.
The image on the right is from the initial upstream" end of the epididymis, the head or caput portion. The epithelium here is of "festooned" appearance, i.e., of varying height.
The epididymis' overall diameter enlarges and the passage through the tubule becomes more open as you progress to the "tail" portion: simultaneously the wall becomes thicker and more muscular. Contraction of the walls and the pressure from "upstream" move sperm through the epididymis. The image below left is from the  "tail" portion: you can easily appreciate the great thickness of the wall, and the fact that it's made of smooth muscle.
"tail" portion: you can easily appreciate the great thickness of the wall, and the fact that it's made of smooth muscle.
 Eventually, when the spermatozoa reach the point where they are physically and physiologically mature, and can be used, they move out of the epididymis and into its extension, the ductus deferens. That organ ultimately connects to the urethra.
Eventually, when the spermatozoa reach the point where they are physically and physiologically mature, and can be used, they move out of the epididymis and into its extension, the ductus deferens. That organ ultimately connects to the urethra.
Ductus Deferens
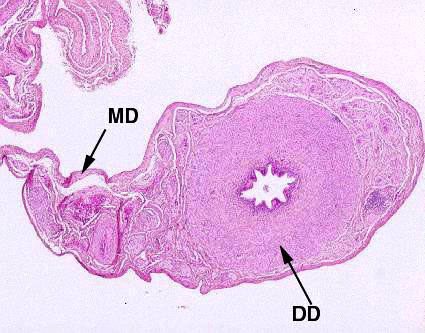 The ductus deferens may be considered the
extra-epididymidal continuation of the cauda of the epididymis. This
portion of the tract is the last stop for sperm, the launching pad for their ultimate
release into the Grand Adventure that awaits them.
The ductus deferens may be considered the
extra-epididymidal continuation of the cauda of the epididymis. This
portion of the tract is the last stop for sperm, the launching pad for their ultimate
release into the Grand Adventure that awaits them.
At left is the ductus deferens, and its supporting CT structure, a fold of the peritoneum, the mesoductus deferens
The ductus deferens is seen on slide 704 in both longitudinal and cross section.
The thickness of the wall in proportion to the cross section is clearly
visible. The mucosal lining is thrown up into folds which run parallel to the
long axis of the organ. The lining epithelium is, as before, a stereociliated
pseudostratified columnar type. The organ as a whole is surrounded by a CT
envelope through which blood vessels and nerves pass.

The presence of so much muscle leads to the questions of what it's for, and how it's made to contract. The smooth muscle cells are served by axons of sympathetic motor neurons; at the moment of climax, the powerful spasm of contraction in the wall of the ductus deferens causes the actual ejaculation of semen. (Now, that's sympathetic inervation!)
Spermatic Cord
Slides 211 and 709 show the spermatic cord, the suspensory structure of
the testis. The cord is a complicated structure, among whose many functions is serving as a route for blood entering and leaving the testis. It carries the principal artery supplying blood to the organ, an
d the veins that drain it (see below); as well as the nervous supply and the ductus
deferens; the whole is bound together with CT.
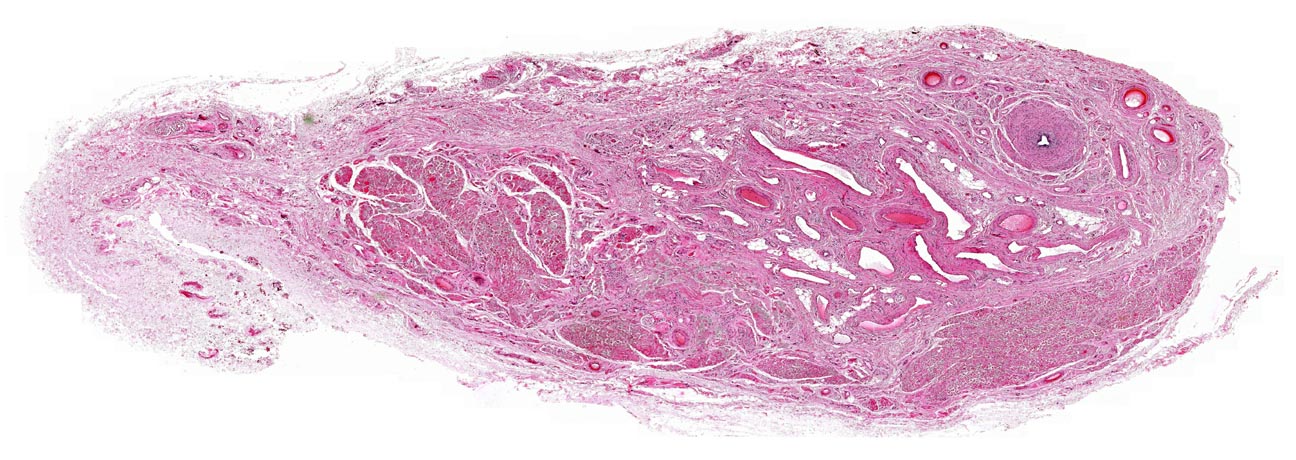
When examining this slide, note the profusion of blood vessels. The veins coming back via the cord form a large plexus, through which the supplying arteries are routed. The arteries and veins thus lie side by side, carrying blood in opposite directions. This intimate physical proximity means that blood moving through the arteries to the testis can be cooled slightly. The opposing flows are arranged into an efficient counter-current heat exchange mechanism; heat moves from the incoming  blood to blood coming out of the testis. Recall that the optimum temperature for spermatogenesis is typically a few degrees below core body temperature: arterial blood is pre-cooled to keep the
temperature of the testes at the ideal point for spermatogenesis, about two
degrees below normal body temperature. The returning blood receives heat, so heat loss from the core is minimized. In either direction, blood encounters a constant temperature gradient and the two flows will never equilibrate.
blood to blood coming out of the testis. Recall that the optimum temperature for spermatogenesis is typically a few degrees below core body temperature: arterial blood is pre-cooled to keep the
temperature of the testes at the ideal point for spermatogenesis, about two
degrees below normal body temperature. The returning blood receives heat, so heat loss from the core is minimized. In either direction, blood encounters a constant temperature gradient and the two flows will never equilibrate.
Don't be surprised to find skeletal muscle (left) as part of the spermatic cord. There's quite a bit, forming the cremaster
muscle, a which can be contracted to
draw the testes up towards the abdomen.
The Penis
 The ductus deferens carries sperm to the urethra, and the sperm are discharged
through that portion of the urethra which passes through the penis. Slide 751 and 707
show examples, cut in cross section. Both of these are from dogs. The labeled specimen at right is human. The urethra is located below the center of the section, and is lined with stratified squamous
epithelium for the last few centimeters: more proximally you would ordinarily see urinary
("transitional") epithelium.
The ductus deferens carries sperm to the urethra, and the sperm are discharged
through that portion of the urethra which passes through the penis. Slide 751 and 707
show examples, cut in cross section. Both of these are from dogs. The labeled specimen at right is human. The urethra is located below the center of the section, and is lined with stratified squamous
epithelium for the last few centimeters: more proximally you would ordinarily see urinary
("transitional") epithelium.
Urethra and Corpora Spongiosa and Cavernosa
Immediately surrounding the urethra are the blood sinuses of the corpus spongiosum, which in this slide are still filled with blood. Dorsally you can see the much larger blood sinuses of the corpus cavernosum, also filled with blood.
These blind vascular channels are involved in the mechanism of erection. A very dense CT wall demarcates the corpus cavernosum, and septa divide it into chambers enclosing the erectile tissue. Careful examination of the blood sinuses in these areas will reveal that they are lined with simple squamous epithelium, as is the rest of the cardiovascular system. The corpus spongiosum around the urethra is also "erectile" tissue, and its job is to keep the urethra patent during ejaculation.
Os Penis
 One structure on slide 707 is worth special notice: the os penis,
found, in domestic animals, only in carnivores.
One structure on slide 707 is worth special notice: the os penis,
found, in domestic animals, only in carnivores.
This triangular shaped bone is located
dorsal to the urethra, and ventral to the corpus cavernosum. As with any bone, its internal
spaces are filled with marrow. There's s a large artery on either side of the os
penis. The longitudinal section on slide 707 demonstrates the surface skin of
this region: stratified squamous, only very slightly keratinized.
Spermatozoa
Having expended so much effort on the means of production and distribution, it's
probably just as well to take some time to look at the final product all this
machinery is designed to make: sperm. Perhaps more than any other cell, the
spermatozoon is best exemplified by a mechanical analogy. In essence, a spermatozoon is little more than a DNA torpedo. Everything that is
not absolutely essential to the overriding mission of "getting there firstest with the mostest" has been stripped away, and the analogy to a torpedo is quite appropriate. There is a power plant, a propulsive mechanism, and—of course—a warhead that does the damage.
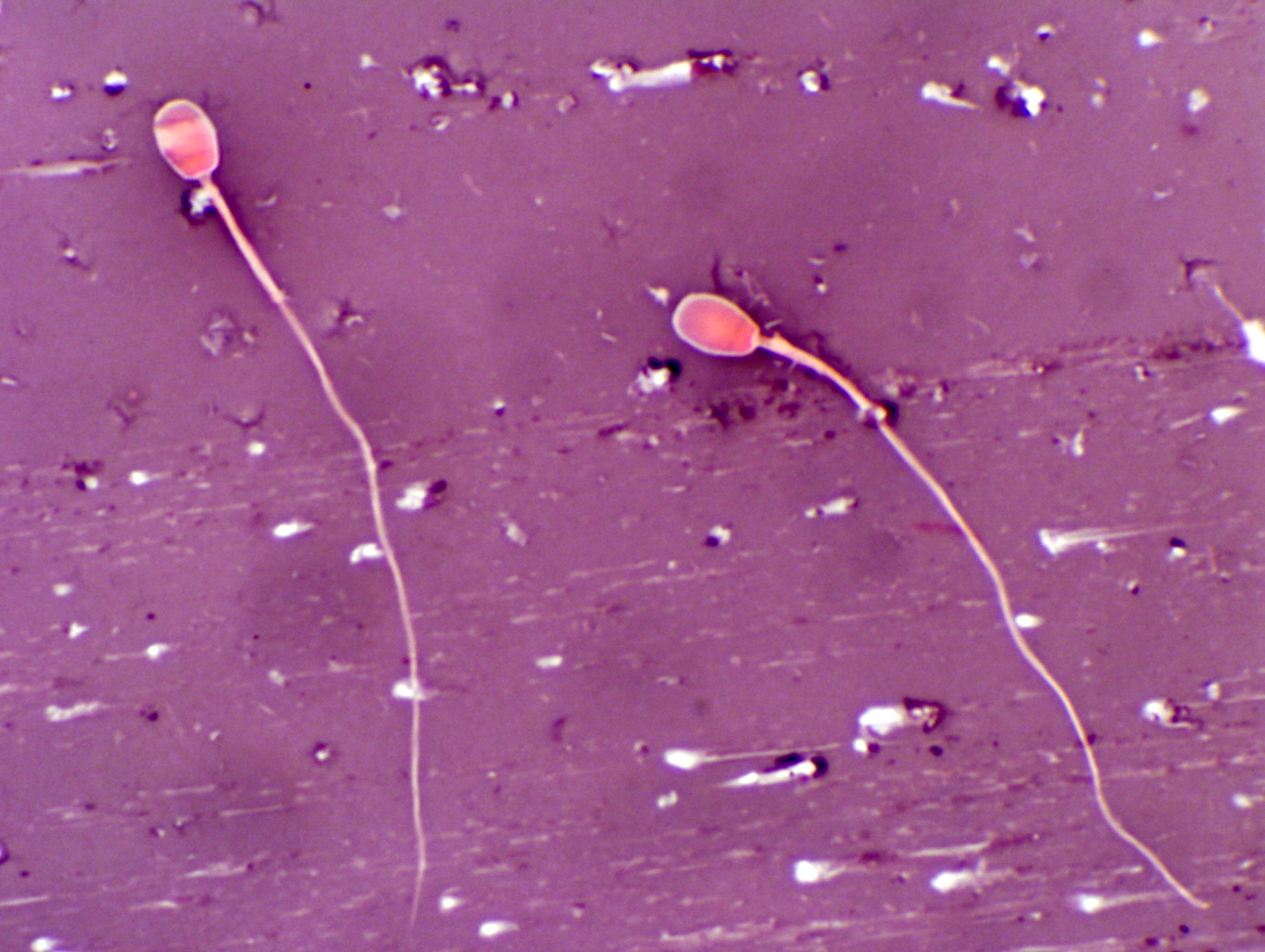
Sperm are produced in almost unbelievable numbers. The typical human male
produces something on the order of 6 million sperm per gram of testicular
tissue; a measly rat about three times that. The all time champion sperm
machine is the pig testis; a boar cranks out about 23 million sperm per gram,
for a total production of 16 billion sperm per day!
You have sperm smears on slides 212 and 934. Slide 212 is from a dog. Select an
area near the edge of the smear where the sperm are not concentrated too
thickly, and you will be able to make out a considerable amount of detail.
(This is one of the rare situations in normal histology where an oil-immersion
lens is useful, so give way to your instincts and use it.)
Head and Acrosome
You should be able to make out on the smears the acrosomal cap in regions where the
staining has been good (there is a considerable degree of variation in this
slide's stain). The acrosome is the detonator, releasing lytic enzymes to dissolve the coats of the oocyte. The acrosomal region covers
about two thirds of this head, and a dark/light transition back near the point
of attachment of the tail marks its boundary.
Connecting Piece, Midpiece, and Endpiece of the Tail
The connecting piece of the tail is the region immediately behind the
head, and is very short. The next part of the tail is the midpiece. This
portion is covered by the mitochondrial sheath, and is noticeably thicker than
the rest of the tail. Behind that is the principal piece. The transition from principal piece to end piece in the tail is not
so marked as from midpiece to principal piece, but you may be able to discern
it. Don't be surprised to see two headed or two tailed sperm; such
abnormalities are fairly common, and in some individual animals they may
represent a very substantial proportion of the number of sperm produced.
Slide 934 is a sperm smear from a rat. The divisions of the tail are
hard to see, but the really interesting thing about this slide is the shape of
the heads. They are described as "scimitar shaped" in texts, a
scimitar being the sort of curved broadswords favored by Barbary
Pirates and Algerine corsairs in the early 19th century. This shape is
characteristic of sperm from all of the rodents of which I am aware. Horses have sperm with heads shaped like broadhead arrows, as do humans. Dogs
have paddle shaped sperm heads. The shapes presumably reflect some sort of
structural adaptation to the challenges faced by a sperm in its quest, but why
there should be so much species variation is anybody's guess.
Accessory Glands of the Male Reproductive System

 Sperm are merely one component of the ejaculate. The male accessory glands produce the rest. In histological sections, most of these look much alike. Of
these glands, by far the most important to clinicians is the
prostate gland, shown here in cross section. The urethra, the opening in the center, is the major landmark by which to recognize this gland. The glandular components take the
form of secretory regions disposed radially around the urethra, shown at upper right.
Sperm are merely one component of the ejaculate. The male accessory glands produce the rest. In histological sections, most of these look much alike. Of
these glands, by far the most important to clinicians is the
prostate gland, shown here in cross section. The urethra, the opening in the center, is the major landmark by which to recognize this gland. The glandular components take the
form of secretory regions disposed radially around the urethra, shown at upper right.
Prostatic tumors (both benign and malignant) are very
common in older dogs (and in men) and they frequently cause difficulty in
urination. As you can
readily appreciate, having all that stuff around the urethra could present
problems if a tumor were present; it would occlude the urethral lumen just as
one might flatten a soda straw. This is another reason why castration is performed on dogs: prostate hyperplasia and tumors are testosterone-dependent and removing the testes prevents them from occurring.
Lab Exercise List