PRE-RECORDED LECTURES FOR THIS EXERCISE
Introduction to the Respiratory System
The Nose
Lower Respiratory Tract
The respiratory system includes all those structures involved in inspiration of air, gas exchange, and expiration of CO2. It also includes the anatomic and histologic structures associated with the chemoreceptor sense of smell. In this exercise we'll examine the histologic features of the nose, the epiglottis and the larynx, the trachea, bronchi, and the lungs.
It's as well to begin at the beginning, at the point where air enters the system: the nose. Of course, most (not all) mammals can breathe through their mouths as well. (The histology of the oral cavity is discussed in another exercise and won't be dealt with again here.)
 On slide 115 you will find a coronal or frontal section of the snout of a mouse.
There are several landmarks on this slide which should help you to get oriented
to where you are. (It helps if you put the slide on the microscope stage in
such a way that the oral cavity is at the bottom of the field; you'll then
have the dorsal and ventral aspects properly positioned.)
On slide 115 you will find a coronal or frontal section of the snout of a mouse.
There are several landmarks on this slide which should help you to get oriented
to where you are. (It helps if you put the slide on the microscope stage in
such a way that the oral cavity is at the bottom of the field; you'll then
have the dorsal and ventral aspects properly positioned.)
This section is viewed from the front: the dorsal side is at the top. The tongue has been cut in cross section, and you can see the connection between the nasal and oral cavities.
The nasal septum (at least this part of it) is made of hyaline cartilage. The vomeronasal organ(s) on either side of the septum are present in this section, as are some of the bony parts of the nose, covered with olfactory epithelium.
Two types of epithelium are present in the nasal cavity, though you can't really see the differences in them at this magnification. The first is typical respiratory epithelium or TRE, the kind of pseudostratified epithelial lining of most of the respiratory tract. The second is the olfactory epithelium, a chemoreceptive type found only in the nose and the vomeronasal organs.
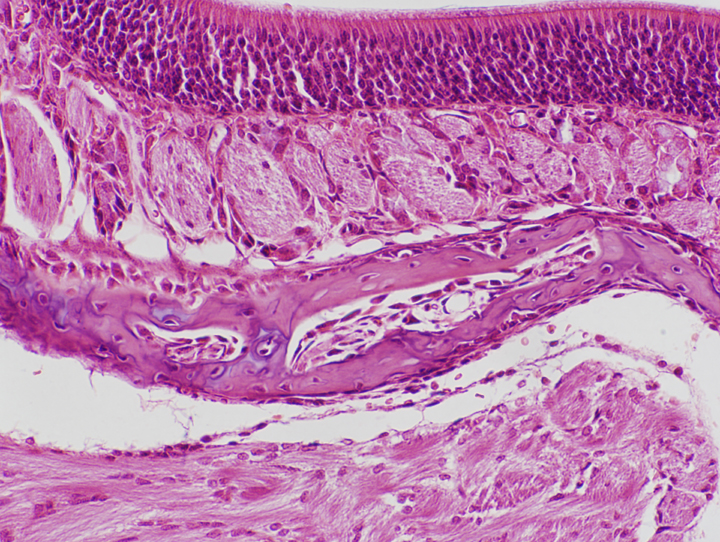
![]() At right and below left are two images of the point of transition between the stratified squamous epithelium of the oral cavity and the pseudostratified epithelium of the nasal cavity. The change-over point is indicated by arrows. The actual border between these two types is fairly sharp, but it's undulating and in a two-dimensional section like this one, hard to pinpoint exactly. The oral epithelium is clearly stratified squamous, and the respiratory epithelium clearly pseudostratified, in both images. In the image at lower left, the oral cavity is below the arrows.
At right and below left are two images of the point of transition between the stratified squamous epithelium of the oral cavity and the pseudostratified epithelium of the nasal cavity. The change-over point is indicated by arrows. The actual border between these two types is fairly sharp, but it's undulating and in a two-dimensional section like this one, hard to pinpoint exactly. The oral epithelium is clearly stratified squamous, and the respiratory epithelium clearly pseudostratified, in both images. In the image at lower left, the oral cavity is below the arrows.
![]()
On either side of the head you'll see two long pieces of bone, running dorsoventrally; these are the bones of the jaw. Lateral to at least one, and possibly both of these, you will see large masses of the muscles which operate the jaw. There should be a salivary gland medial to the jawbone on either side, and there are very large nerve processes, cut in cross section, running "through" the slide from above to below the plane of the section.
Once you've figured out which way is up, examine the nasal cavity, the open space dorsal to the oral cavity, and separated from it by a bony plate running parallel to the plane of the section.

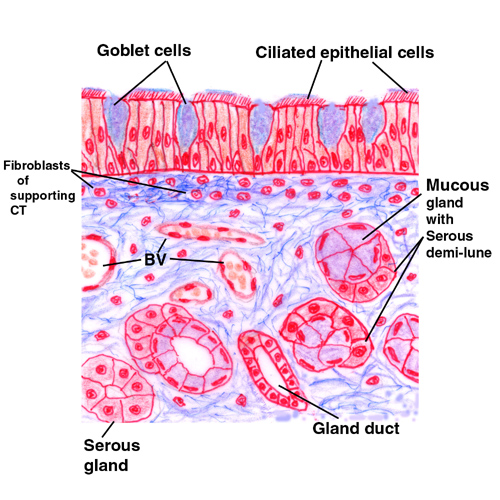 The nose is the first place where you'll encounter typical respiratory epithelium,
usually abbreviated as "TRE." This term refers to a sheet of ciliated
pseudostratified columnar epithelium with goblet cells scattered
throughout. You'll find TRE throughout the length of the air passages of the
respiratory tract. In the nose it lines most of the cavity, except for the for
the chemosensory area. In the deeper part of the system it disappears before
you get to the very end of the smallest airways. But it's the characteristic
lining epithelium for the bulk of the respiratory system, from the nose all the way to small bronchioles. In slide 115, look for small open areas dorsal to the oral cavity and you'll find nice examples
of TRE, where the ciliary fringe is very obvious.
The nose is the first place where you'll encounter typical respiratory epithelium,
usually abbreviated as "TRE." This term refers to a sheet of ciliated
pseudostratified columnar epithelium with goblet cells scattered
throughout. You'll find TRE throughout the length of the air passages of the
respiratory tract. In the nose it lines most of the cavity, except for the for
the chemosensory area. In the deeper part of the system it disappears before
you get to the very end of the smallest airways. But it's the characteristic
lining epithelium for the bulk of the respiratory system, from the nose all the way to small bronchioles. In slide 115, look for small open areas dorsal to the oral cavity and you'll find nice examples
of TRE, where the ciliary fringe is very obvious.
TRE is a pseudostratified, ciliated, columnar epithelial sheet liberally interspersed with goblet cells. It rests on a CT lamina propria containing a large collection of glands and blood vessels. One function of the nose is to warm incoming air, and to trap some (by no means all) of the dirt and bacterial particles that are inhaled. The blood vessels underneath the epithelium serve to transfer heat to the incoming air flow, so that the humidity of the lung can be maintained at saturation.
 The serous and mucous secretions produced by the nasal glands underlying the epithelium (and the goblet cells embedded in the epithelial sheet) serve to catch and hold any dirt particles and bacteria that are inhaled. Keeping the surface moist provides a fluid sheet that traps things; the ciliated cells then can move the moist sheet as a whole, the beating of the cilia being unidirectional. This is much less important in the nasal cavity than it is in the deeper parts of the respiratory tract, however.
The serous and mucous secretions produced by the nasal glands underlying the epithelium (and the goblet cells embedded in the epithelial sheet) serve to catch and hold any dirt particles and bacteria that are inhaled. Keeping the surface moist provides a fluid sheet that traps things; the ciliated cells then can move the moist sheet as a whole, the beating of the cilia being unidirectional. This is much less important in the nasal cavity than it is in the deeper parts of the respiratory tract, however.
Olfactory Epithelium
The nasal cavity has two regions of pseudostratified epithelium, the respiratory area (which we've looked at) and the olfactory epithelium which replaces it in some regions. Towards the dorsal part of nasal cavity you'll find spiraling bony plates (the nasal turbinates) covered with a high pseudostratified columnar epithelium. This is the portion of the nasal cavity specialized for chemoreception.
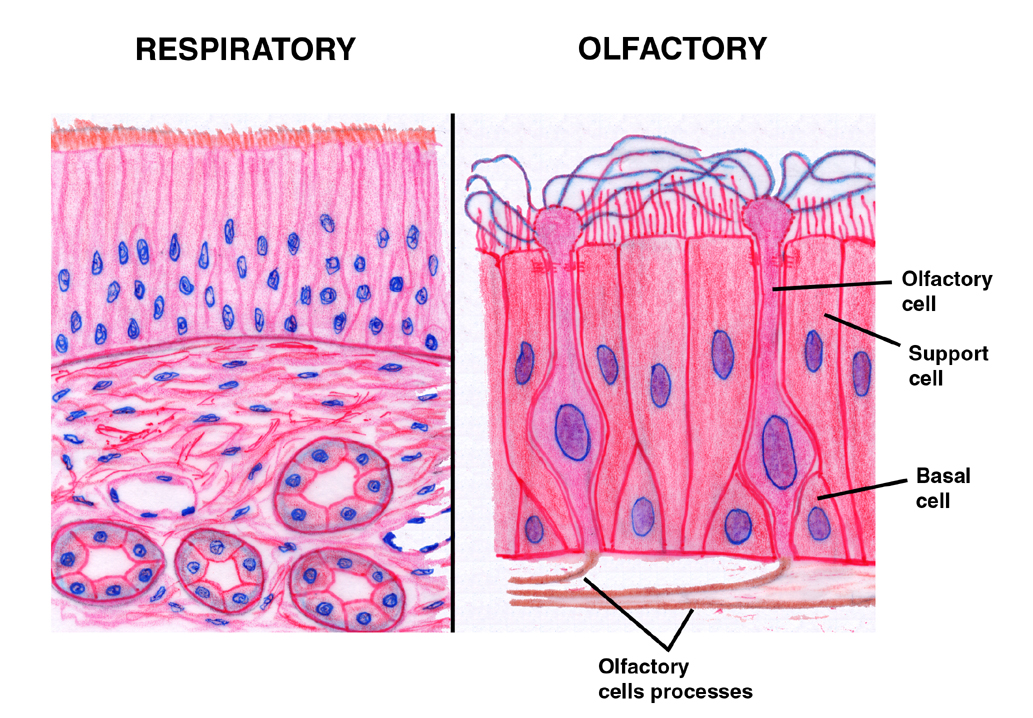 This olfactory
epithelium is also ciliated, but its function isn't to trap dust and
microbes, as is that of the TRE. Olfactory epithelium is much higher than TRE, which is related to its chemoreceptive function. Both TRE
This olfactory
epithelium is also ciliated, but its function isn't to trap dust and
microbes, as is that of the TRE. Olfactory epithelium is much higher than TRE, which is related to its chemoreceptive function. Both TRE  and olfactory
epithelium are on the slide, so you can compare them directly. A schematic comparison is shown at left. At left is a scanning electron micrograph of the olfactory area: the cilia are obvious, but far less numerous than in the respiratory regions.
and olfactory
epithelium are on the slide, so you can compare them directly. A schematic comparison is shown at left. At left is a scanning electron micrograph of the olfactory area: the cilia are obvious, but far less numerous than in the respiratory regions.
Olfactory epithelium is found on the turbinates and also in a specialized area, the vomeronasal organ. It's a very high pseudostratified columnar type. The cilia are quite easily made out at the free surface but typically there are few or no goblet cells in the olfactory areas. Three cell types are present, only one of which is chemoreceptive.
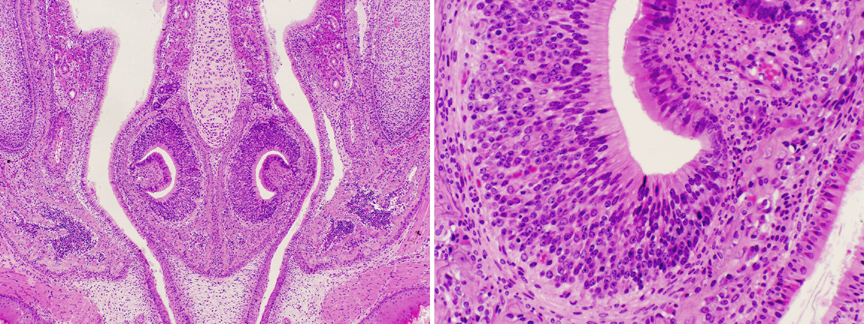
This image nicely illustrates the vomeronasal organ, and its areas of chemoreceptive olfactory epithelium. The VNO apparently has special connections to the olfactory processing area that is related to perception of pheromones, especially those concerned with reproduction. Its placement in the air stream is such that it gets maximum flow, and the architecture allows for slowing of the moving air to increase sensitivity. The olfactory epithelium in this structure is very high pseudo-stratified, with numerous specialized olfactory cells (see below).

The arrangement on the dorsal portion of the nose, the turbinate bones, and the upper part of the nasal septum is similar; it's shown above.

 The olfactory cells themselves (OC) are transducers of chemical sensations into neural signals. The mechanism of smell is still very obscure, but it is known that the plasma membrane of the olfactory cells is the actual site of chemoreception. One of these cells is shown here at about 1000x. It's a tall cell that reaches both the free surface and the basement membrane. Olfactory cells are really a form of bipolar neuron: as with any other neuron, they're so specialized they need a set of supporting cells (SC) to tend to their needs. The supporting cells and the basal cells (BC) can be thought of as quasi-glial elements; they perform the same functions of protection. Not visible in this light micrograph, but demonstrable in an EM are occluding junctions that seal the apex of the olfactory cell to its supporting cells and to the basal cells. Thus the body of the olfactory cell is effectively isolated from outside influences and only the modified cilia that are the actual site of transduction, and the axon leading from the base, are "exposed."
The olfactory cells themselves (OC) are transducers of chemical sensations into neural signals. The mechanism of smell is still very obscure, but it is known that the plasma membrane of the olfactory cells is the actual site of chemoreception. One of these cells is shown here at about 1000x. It's a tall cell that reaches both the free surface and the basement membrane. Olfactory cells are really a form of bipolar neuron: as with any other neuron, they're so specialized they need a set of supporting cells (SC) to tend to their needs. The supporting cells and the basal cells (BC) can be thought of as quasi-glial elements; they perform the same functions of protection. Not visible in this light micrograph, but demonstrable in an EM are occluding junctions that seal the apex of the olfactory cell to its supporting cells and to the basal cells. Thus the body of the olfactory cell is effectively isolated from outside influences and only the modified cilia that are the actual site of transduction, and the axon leading from the base, are "exposed."
The axons of the olfactory cells are bundled together to form the olfactory nerve, which is then routed through the overlying bone of the nose into the olfactory lobe of the brain. In the image at left, the perforation through the bone of a few of these fiber bundles is shown. The low-power image at right  shows one of the two olfactory nerves after it's actually formed a root, at the point where it enters the olfactory lobe proper. Note the numerous layers of neurons in the lobe, which are devoted to processing and routing the incoming signals from thousands of nerve fibers.
shows one of the two olfactory nerves after it's actually formed a root, at the point where it enters the olfactory lobe proper. Note the numerous layers of neurons in the lobe, which are devoted to processing and routing the incoming signals from thousands of nerve fibers.
Because they're neurons, olfactory cells are "excitable," i.e., capable of responding to changes in the environment around them. Olfaction is believed to occur when molecules of a scent (an "olfactant") bind to appropriate membrane locations. In so doing they modify the plasma membrane's charge characteristics, which triggers a signal; this signal, essentially an action potential, is routed via an afferent fiber to a pseudo-unipolar neuron in one of the cranial nerve ganglia; from thence it's relayed to the brain's olfactory signal-processing location, the olfactory lobe. Further signal integration takes place within the brain by obscure pathways. Animals with a keen sense of smell typically have a great many olfactory cells present, and very large olfactory lobes to process the input.
 If the olfactory cells' binding sites aren't cleared of the materials bound to
them, after a while they no longer transmit a signal, a phenomenon called "olfactory fatigue." After you've been in a barn for a few minutes you "don't
smell it anymore." Input from the chemoreception
area is reduced partly due to saturation of available odorant binding sites; and partly because and your brain is filtering out what signals get through as "normal" and "harmless."
If the olfactory cells' binding sites aren't cleared of the materials bound to
them, after a while they no longer transmit a signal, a phenomenon called "olfactory fatigue." After you've been in a barn for a few minutes you "don't
smell it anymore." Input from the chemoreception
area is reduced partly due to saturation of available odorant binding sites; and partly because and your brain is filtering out what signals get through as "normal" and "harmless."
Obviously, this is an inconvenient—not to say dangerous—situation if you're an animal that relies on its nose for warning of danger approaching. Hence, a mechanism to wash the olfactory receptors clean, and to make them responsive again, exists.
As in the respiratory region, there are serous glands in the lamina propria under the olfactory region. Secretions from these clean the sensory structures and ready them for reuse. If you own a dog, watch him the next time he comes to some especially ripe and disgusting thing he wants to smell. He will inhale deeply, and then his nose starts to drip. What's happening is that these olfactory glands are kicking into high gear and producing copious secretions to keep that's dog's sense of smell at its peak of perfection, lest he miss something—or someone—really worth smelling.
 Epiglottis and Larynx
Epiglottis and Larynx  The entry into the lower part of the respiratory system is guarded by the epiglottis,
cut in longitudinal section on slide 619. This is a flap of elastic
cartilage, as you can easily see, and it's covered on all sides by
epithelium. Note the glands in the lamina propria on this slide. Glands are found throughout
most of the airways, and their secretions help to trap dust and debris so that the beating of the ciliated cells can bring it up to
the pharynx, where it is swallowed or spit out.
The entry into the lower part of the respiratory system is guarded by the epiglottis,
cut in longitudinal section on slide 619. This is a flap of elastic
cartilage, as you can easily see, and it's covered on all sides by
epithelium. Note the glands in the lamina propria on this slide. Glands are found throughout
most of the airways, and their secretions help to trap dust and debris so that the beating of the ciliated cells can bring it up to
the pharynx, where it is swallowed or spit out.
Slide 620 is a section of a larynx, the cartilaginous box into which the epiglottis opens. This is shown at right, cut longitudinally. The pharyngeal end of the larynx is at the top of this low-power view. The larynx is a box constructed of plates of hyaline and some elastic cartilage, articulated together with CT. The muscles controlling the shape of the box insert onto the cartilage. On the oral cavity side of the glottis, the lining of the system is stratified squamous, and several laryngeal tonsils can be seen.  Once past the entrance there's an abrupt transition from stratified squamous epithelium of the oral cavity to respiratory epithelium.
Once past the entrance there's an abrupt transition from stratified squamous epithelium of the oral cavity to respiratory epithelium.
At higher magnification the details of the wall are visible. The muscles inserting onto the outside wall can change the shape of the box, and in so doing change the tension and vibration frequencies of the vocal folds. Air being moved through from the depths of the respiratory system causes the vibration to occur. Overtones, harmonics, and other sounds are added by the echoes produced in the chest cavity, the tongue, the teeth, and the oral cavity. Phonation involves distortion of the shape so that the vocal folds at the cranial end vibrate at different rates and produce different tones. This is exactly the same principle by which sounds are produced in that classic piece of high-tech wizardry, the Whoopee Cushion.
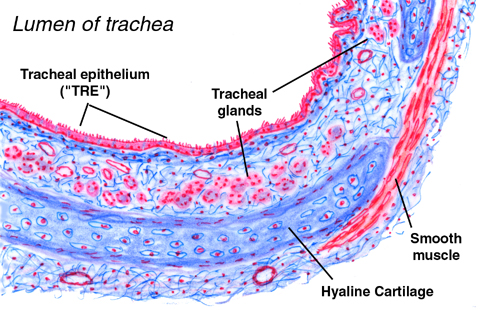 The larynx opens at its lower end into the trachea, seen on slides 26 and
263. On these slides the organ is cut in cross section, but only part of the
entire circumference of the trachea is included. A schematic diagram of the wall is shown at left.
The larynx opens at its lower end into the trachea, seen on slides 26 and
263. On these slides the organ is cut in cross section, but only part of the
entire circumference of the trachea is included. A schematic diagram of the wall is shown at left. The trachea is constructed like the vent hose on a clothes dryer: stiffening C-shaped rings hold it open and flexible segments between the rings contain the air flowing through it. It is lightweight, flexible, and resists compression.
The tracheal rings are composed of hyaline cartilage, not the elastic cartilage of the epiglottis. The respiratory epithelium on these slides is well preserved, and you should be able to make out the cilia quite easily. There are few glands in the lamina propria here. While they aren't too obvious on slide 26, they really stand out on slide 263, which stained with PAS. There are also some blood vessels present in the lamina propria.

Here are two views of the wall, the one on the left stained with H&E, the one on the right with PAS. These should be compared with the schematic. The tracheal lumen, like most of the respiratory tract, is lined with TRE. The goblet cells in the epithelium itself, and the large glands in the sub-epithelial CT, connected to the surface by ducts, contribute to the dust-trap effect. The tracheal glands, located in the lamina propria, are strongly PAS-positive, as you would expect, since they make a carbohydrate-rich secretion. The cartilage itself is PAS-positive thanks to its matrix material. The PAS stain also reacts to the mucous sheet on top of the lining epithelium.
The cilia in the TRE all beat in a coordinated manner to move the sheet of mucus upwards. Mucus is produced continuously, so if it weren't removed, it would accumulate in deeper regions and the animal would shortly drown. Moving the mucus layer up and out takes the trapped debris with it; at the top, it's either swallowed or spit out. We featherless bipeds call these respective actions "clearing the throat" and...ahem..."hawking an oyster."
Bronchi & Bronchioles
The trachea bifurcates on entering the substance of the lung, and further subdivisions occur, several orders of them. Each primary branch, or primary bronchus (bronchi) supplies air to the lung on each side of the chest (cattle and pigs have an extra primary bronchus). For purposes of discussion, we can define a "bronchus" as: an airway whose walls are reinforced by the presence of cartilage plates.
The structure of bronchi is similar to that of the trachea: flexible tubes with stiffening walls of hyaline cartilage. However, once in the substance of the lungs, the ring shaped cartilage support of the trachea gives way to cartilage plates. At first these overlap considerably but as the air passages decrease in size, there's a corresponding decrease in the size of the cartilage plates, and an increase in the smooth muscle component of the wall. Sooner or later, the last bits of cartilage disappear and the wall is reinforced only with smooth muscle. At that point the airway is small enough and the muscle tonus great enough to hold the airway patent.
For our purposes the distinction between the various orders of airways isn't important, so we'll recognize only two classes: bronchi, and the bronchioles, those airways whose walls contain no cartilage. In the discussion below, if cartilage is present, it's a "bronchus," but if cartilage is absent the airway is a "bronchiole." Neither has a gas exchange function: that occurs at an even deeper level of the system.
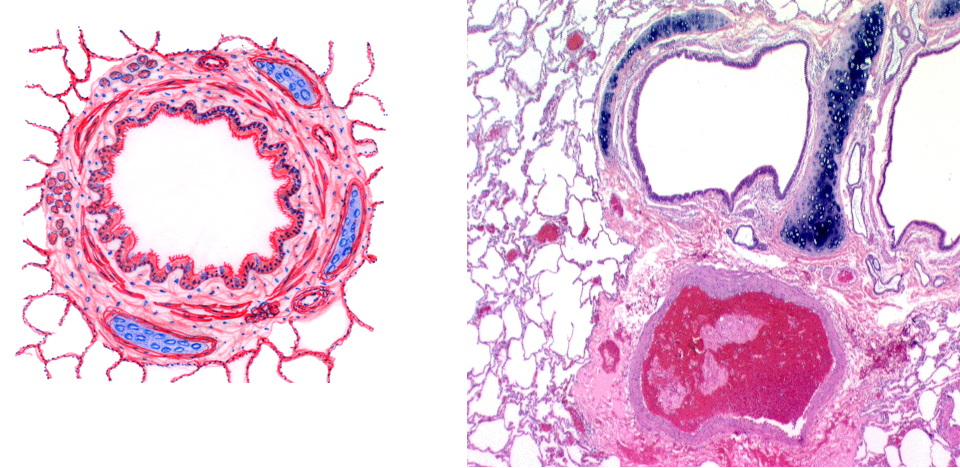
Here's a schematic diagram of a medium-sized bronchus and a real one for comparison. These vessels are cut in cross section. The cartilage reinforcement is pretty obvious. In the wall of the bronchi you'll also find strands of smooth muscle, which increases in proportion and continuity as the vessel decreases in size. In this example there muscle is still a minor component of the wall. Note also the bronchial glands in the lamina propria: these serve the same moistening and dust-trap functions as the glands in the trachea do. The lining of all bronchi is TRE. In the image on the right you can also see one of the large arteries that run alongside the bronchi: these are branches of the pulmonary artery. Numerous smaller blood vessels are present in among the profiles of the gas exchange areas, as well.

Here's a rather larger bronchus, but cut in longitudinal (or partially oblique) section; a smaller branch is given off at the right side. Notice that in cross section, the lining of the bronchus is plicated (thrown up in folds). this is an adaptation to expansion as the lungs inflate. In longitudinal sections, these plications give the bronchus an odd "scalloped" appearance. The reinforcement of  the wall is very obvious. This bronchus is big enough that the plates overlap quite a bit, forming an articulated wall. Connective tissue (a mixture of collagenous and elastic, that is continuous with the perichondrium of the plates) fills the gaps between the collagen plates. There are bundles of smooth muscle in the lamina propria between the cartilage and the lining of TRE, and some bronchial glands are present. The blood vessel alongside the bronchus is part of the pulmonary arterial system.
the wall is very obvious. This bronchus is big enough that the plates overlap quite a bit, forming an articulated wall. Connective tissue (a mixture of collagenous and elastic, that is continuous with the perichondrium of the plates) fills the gaps between the collagen plates. There are bundles of smooth muscle in the lamina propria between the cartilage and the lining of TRE, and some bronchial glands are present. The blood vessel alongside the bronchus is part of the pulmonary arterial system.
As the bronchi diminish in size, the proportion of cartilage in the wall decreases and the proportion of smooth muscle (SM) increases. The edges of the plates are spaced apart from each other and the "gaps" are filled with CT and muscle.
In the image at left you see a somewhat smaller bronchus cut in oblique section. This section is from slide 100; it's stained with the Masson's stain. The cartilaginous plates (CP) have been reduced considerably in size here, to the point where they no longer overlap. The reinforcement of smooth muscle (stained red-brown with this method) constitutes a much larger and more significant section of the wall. Again, there's an artery running below it.
 Eventually, in the very smallest airways there will be no cartilage left. The wall reinforcement will be composed entirely smooth muscle. This absence of cartilage is the hallmark, the principal structural difference between a "bronchus" and a "bronchiole" as we've defined the two. You see this situation diagrammed and in an actual specimen, here. The cartilage is needed in large bronchi because their tube diameter is comparatively large. To prevent collapse as the lungs expand, it's necessary to stiffen the wall with the cartilage plates. But when the airway gets below a diameter of about 0.5 mm, the smooth muscle in the wall has sufficient tone to resist collapse. There's no point in expending metabolic energy to make reinforcement that isn't required.
Eventually, in the very smallest airways there will be no cartilage left. The wall reinforcement will be composed entirely smooth muscle. This absence of cartilage is the hallmark, the principal structural difference between a "bronchus" and a "bronchiole" as we've defined the two. You see this situation diagrammed and in an actual specimen, here. The cartilage is needed in large bronchi because their tube diameter is comparatively large. To prevent collapse as the lungs expand, it's necessary to stiffen the wall with the cartilage plates. But when the airway gets below a diameter of about 0.5 mm, the smooth muscle in the wall has sufficient tone to resist collapse. There's no point in expending metabolic energy to make reinforcement that isn't required.
Bronchi proper can be as small as 0.5 mm diameter and still retain their lining of TRE, but when they get much smaller than that, some changes occur. First, the goblet cells disappear, and long before the bronchioles have got down to the half-millimeter range the glands have disappeared. Eventually there is an epithelial transition from the pseudostratified TRE, first to simple columnar, then to a low cuboidal type. Along the way ciliation disappears, but this doesn't happen until after the goblet cells are lost. Why? Because otherwise mucus would be produced that couldn't be cleared. hence secretion ends before ciliation does.
We have looked so far only at the airways, and now it's time to see where the actual gas exchange occurs, in the lungs. The description given below applies to mammalian lungs. Bird lungs are very different and will be dealt with separately.
Mammals are "negative pressure breathers." Mammalian lungs are always partially inflated. This is accomplished by maintaining the lungs inside an airtight sac, rather like a plastic bag, inside the chest; the pressure in the space between the sac and the chest wall is lower than atmospheric pressure. If something causes this pressure differential to be removed (for example, a penetrating or perforating trauma to the chest wall) the elasticity of the lungs will cause them to collapse. This condition is a pneumothorax, which if bilateral, is very serious because the animal will die quickly from anoxia.
Inhalation in mammals involves contraction of the muscular diaphragm and expansion of the chest wall. This increases the volume of the chest, and hence the pressure differential between it and atmosphere, and the lungs inflate. Relaxation of the muscles causes the chest to decrease in size, the diaphragm to relax, and the exhalation of the cycle occurs.
For this part of the exercise, we'll use slides 100, 111, and 559.
Slide 111 is a piece of lung. On the outside surface of the section, you should be able to make out a thin layer of simple squamous epithelium overlying a very delicate CT. This is the visceral pleura, which corresponds in nature and origins to the peritoneum of the abdominal cavity. It's formed from the mesoderm of the primitive pleural cavity into which the lung bud expands during organogenesis.
 Here's a nice example of lung as it's supposed to look, from slide 100. This is stained with the Masson's routine. The walls of the gas exchange areas, the alveoli are "barely there"—they consist of a very tenuous covering of squamous cells (Type I pneumocytes) over capillaries. A lung that looks like one you see here is fully functional, and exchange of gases across the plasma membranes of the pneumocytes and the underlying blood vessels is unhindered.
Here's a nice example of lung as it's supposed to look, from slide 100. This is stained with the Masson's routine. The walls of the gas exchange areas, the alveoli are "barely there"—they consist of a very tenuous covering of squamous cells (Type I pneumocytes) over capillaries. A lung that looks like one you see here is fully functional, and exchange of gases across the plasma membranes of the pneumocytes and the underlying blood vessels is unhindered.
The functional unit of the lung is the alveolus. Alveoli are blind cul-de-sacs, butted back-to-back against each other. The wall of the alveolus is made of the thin Type I pneumocytes overlying capillaries.
 The nature of alveoli can be hard to grasp from a section but you can refer to the sketch below for a better understanding of how the alveoli are built and how they relate to each other. This drawing shows four individual alveoli, forming a group, an alveolar sac. The sac is at the end of the respiratory tree, i.e., at the end of the smallest bronchiole serving this region. On the left side is show the external surface of the alveolar sac; on the right side it has been drawn as if sectioned, so that the interior is exposed and the connective tissue that supports the alveolar walls is exposed.
The nature of alveoli can be hard to grasp from a section but you can refer to the sketch below for a better understanding of how the alveoli are built and how they relate to each other. This drawing shows four individual alveoli, forming a group, an alveolar sac. The sac is at the end of the respiratory tree, i.e., at the end of the smallest bronchiole serving this region. On the left side is show the external surface of the alveolar sac; on the right side it has been drawn as if sectioned, so that the interior is exposed and the connective tissue that supports the alveolar walls is exposed.
Individual alveoli often communicate back-to-back with each other via alveolar pores. This allows more complete exchange of the volume of air in an individual alveolus, an important functional consideration. The bronchioles are lined with cuboidal epithelium (Type I pneumocytes) and the alveoli with a thin squamous type that covers the capillaries in the connective tissue of the alveolar walls. Type II pneumocytes are scattered here and there in the alveolar walls, wedged in between the Type I cells and bulging outward into the lumen of the alveolus. Free macrophages (not shown here) may be found in the alveoli or wandering around on the surface; or they may be "in transit" between the CT or the alveolar lumen. Macrophages also use the alveolar pores to get from one alveolus to another.
Using slides 100, 559, and 111 you should now move on to identify specific respiratory structures. All these slides provide very good, open preparations, and there should be places where the end of the purely conductive portions of the lung empties into the gas exchange areas.
Gas exchange doesn't occur in bronchioles. The gas exchange areas are the alveolar ducts, alveolar sacs, and individual alveoli. (The "respiratory bronchioles" discussed in the text—which deals with humans exclusively—probably will not be identifiable in these slides, as they are scarce in most domestic animals and totally absent from some.)
The lung in a sense is really part of the circulatory system. It's essentially an immense capillary plexus, and the blood vessels that run through it are overlain by a squamous epithelium so thin its very existence was a matter of debate until the electron microscope settled the question. At one time it was believed that the lung capillaries were actually "naked," i.e., that the blood in them was separated from the air in the lung only by the capillary endothelial wall. This isn't the case, however; it's now well established that the actual physical barrier to gas exchange consists of the capillary endothelium, its basal lamina, the basal lamina of the alveolar cell, and the thin sheet of alveolar epithelium atop that. It is the alveolar cells which are exposed to the air.
 Individual alveoli should present no problems of identification. The
alveolus is as tenuous a structure as one can imagine, and in these
preparations will be seen cut through "edge-on" only. If you use oil
immersion, especially on slides 100 and 559, you should be able to make out some
details of the structure of the alveolar wall and the relationships of blood
vessels and the pulmonary epithelium.
Individual alveoli should present no problems of identification. The
alveolus is as tenuous a structure as one can imagine, and in these
preparations will be seen cut through "edge-on" only. If you use oil
immersion, especially on slides 100 and 559, you should be able to make out some
details of the structure of the alveolar wall and the relationships of blood
vessels and the pulmonary epithelium.
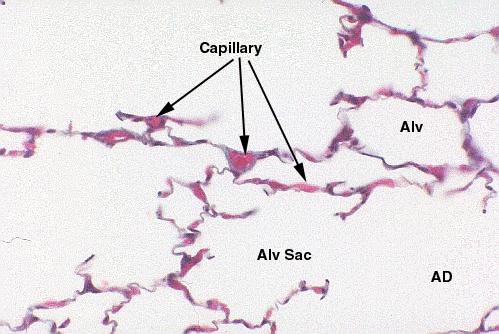 In these two images you see parts of several alveoli. The capillaries running between two
adjacent alveoli are gorged with blood, and pretty well defined by the
erythrocytes in them. Any two neighboring
alveoli are separated from each other by a blood vessel, which is therefore exposed to the air on either side, facilitating gas exchange. There are of course
numerous larger blood vessels, which accompany the airways into the deepest
parts of the lung. These are branches of the pulmonary artery, by which the
lung's substance is perfused.
In these two images you see parts of several alveoli. The capillaries running between two
adjacent alveoli are gorged with blood, and pretty well defined by the
erythrocytes in them. Any two neighboring
alveoli are separated from each other by a blood vessel, which is therefore exposed to the air on either side, facilitating gas exchange. There are of course
numerous larger blood vessels, which accompany the airways into the deepest
parts of the lung. These are branches of the pulmonary artery, by which the
lung's substance is perfused.  The actual gas exchange barrier consists of the Type I pneumocytes, their basement membrane, the basement membrane of the capillary underneath, and the capillary epithelium. The sketch at right should be compared with the image below, to get an idea of how tenuous this barrier really is.
The actual gas exchange barrier consists of the Type I pneumocytes, their basement membrane, the basement membrane of the capillary underneath, and the capillary epithelium. The sketch at right should be compared with the image below, to get an idea of how tenuous this barrier really is.
 The alveoli and alveolar sacs are the dead end of the system. They arise off of alveolar ducts, regions that simultaneously have a gas exchange function and serve as a final airway. The alveolar duct can be thought of as a "place," not a "thing." If you can imagine a tube whose wall is composed of halved soap
bubbles, look for places where the smooth muscle fibers at the
"seams" between bubbles are prominent, and where such
"seams" are lined up. This will be the most reliable way to identify
the alveolar ducts. In the image at left the deepest level of the bronchioles (B) opens into an alveolar duct (AD) which in turn leads to several alveolar sacs (AS). The latter are blind cul-de-sacs of 3 to 5 alveoli.
The alveoli and alveolar sacs are the dead end of the system. They arise off of alveolar ducts, regions that simultaneously have a gas exchange function and serve as a final airway. The alveolar duct can be thought of as a "place," not a "thing." If you can imagine a tube whose wall is composed of halved soap
bubbles, look for places where the smooth muscle fibers at the
"seams" between bubbles are prominent, and where such
"seams" are lined up. This will be the most reliable way to identify
the alveolar ducts. In the image at left the deepest level of the bronchioles (B) opens into an alveolar duct (AD) which in turn leads to several alveolar sacs (AS). The latter are blind cul-de-sacs of 3 to 5 alveoli.
 These are the thin squamous cells overlying the capillaries, that have been mentioned. Their nuclei are all you'll be able to make out. In this image a nucleus is labeled, as are some erythrocytes in capillaries. The capillaries are in fact overlain by the cytoplasm of adjacent squamous Type I cells, but there's so little of it isn't visible.
These are the thin squamous cells overlying the capillaries, that have been mentioned. Their nuclei are all you'll be able to make out. In this image a nucleus is labeled, as are some erythrocytes in capillaries. The capillaries are in fact overlain by the cytoplasm of adjacent squamous Type I cells, but there's so little of it isn't visible.
We have described the thin, squamous cells of the alveolar wall. Two more cell types remain to be described. One is the septal cell or great alveolar cell, sometimes also referred to as a Type 2 pneumocyte. These are found in the walls of the alveoli, often between two adjacent ones. They are low cuboidal type cells, which stick out a bit from the flattened septal epithelium. The Type II cell is part of the alveolar wall: it can usually be identified by the fact that it bulges out and it's usually located at the intersection of two or more alveoli back to back. For this reason it's sometimes called the "septal cell" and its large size has also caused it to be term the "great alveolar cell."

These cells have two functions. First, they're a reserve cell which may as needed differentiate into new squamous cells to repair damage. Second, they produce surfactant, a lipid rich material that coats the inside of the alveoli. Surfactant reduces the surface tension of the thin aqueous film that covers the cells. It's is necessary to keep the very small alveoli from collapsing under the surface tension of their water film, which is considerably greater than their mechanical strength. Surfactant intercalates itself between water molecules and lessens their intermolecular attractions.
 You may also notice a large number of free cells in the lumen of the alveoli.
These are macrophages, which are mobile phagocytic scavengers. They're
very numerous in mammalian lung: their job is to keep the surface of the lung
as sterile as possible by scarfing up bacteria and any other small bits of
debris which make it past the filters and dust traps in the upper regions. In
disease states their numbers increase greatly as monocytes migrate to the lungs
and differentiate into macrophages.
You may also notice a large number of free cells in the lumen of the alveoli.
These are macrophages, which are mobile phagocytic scavengers. They're
very numerous in mammalian lung: their job is to keep the surface of the lung
as sterile as possible by scarfing up bacteria and any other small bits of
debris which make it past the filters and dust traps in the upper regions. In
disease states their numbers increase greatly as monocytes migrate to the lungs
and differentiate into macrophages.
Two are shown at left, indicated by the arrow. They're in the lumen of the alveoli, having reached there by squeezing between Type I pneumocytes, after traveling through the CT septations between the alveoli. Macrophages are a CT cell type, and when fully differentiated they have amoeboid motility, allowing them to crawl from one place to another.
 Macrophages can be quite active, as the image to the right indicates. This is a section from a monkey lung, Four macrophages are pretty obvious, and there's another one on the right edge of the field that's free in the lumen. The black material in these cells is likely environmental contaminants, probably soot.
Macrophages can be quite active, as the image to the right indicates. This is a section from a monkey lung, Four macrophages are pretty obvious, and there's another one on the right edge of the field that's free in the lumen. The black material in these cells is likely environmental contaminants, probably soot.
Isolated macrophage activity like this usually indicates some local problem that eventually gets resolved; the lung of this animal was otherwise in good shape. The large amount of blood in the nearby capillaries is probably related to whatever's going on here; blood flow into the area, increased in response to chemical signals, brings in more macrophages and other immune-system cells if needed.
Macrophages arise from precursor cells in the bone marrow. They're released into the blood, in the immature circulating form we know as the monocyte. The monocyte/macrophage-to-be uses the blood strictly as a transport medium, however. Once it arrives where it's going to be used, it leaves the blood and completes its differentiation. This process of recruitment, transport, and differentiation is especially marked in the case of lung infections: a normal lung may have a few million macrophages patrolling its surface, but an infection will cause the number to skyrocket into the tens or hundreds of millions to deal with the invasion.
![]()