 Cresyl Violet & Luxol Fast Blue
Cresyl Violet & Luxol Fast Blue SLIDES USED IN THIS EXERCISE: 552, 540, 541, VM34, VM25, MS_l02_097, VM39, VM36, 550, 539, 1210, 270/270A, 538, 2-28, 508, 509
THERE ARE PRE-RECORDED LECTURES FOR THIS EXERCISE
Please go to the listing page for these to download and view them
Introduction to Nervous Tissue
Cells of Nervous Tissue
Nerves
Nervous System Organs
In this laboratory exercise you will see slides prepared using stains somewhat different to those used in previous exercises. While H&E is useful as a general structural stain in most tissues, the high lipid content of nervous tissue makes it less suited to H&E than most others. Consequently, numerous stains have been developed which take advantage of the chemical properties of the lipids in neural tissue to highlight structural features.
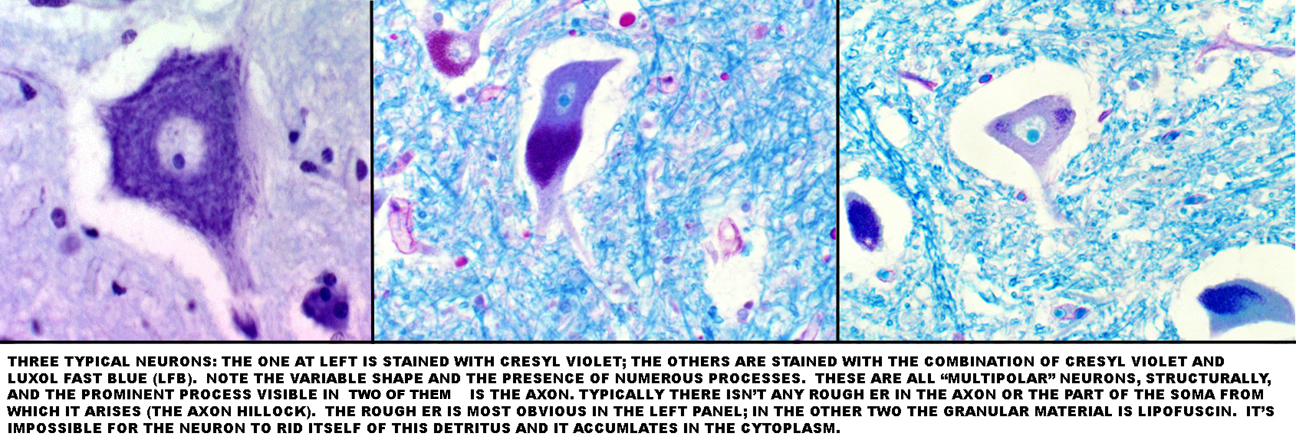
Cresyl Violet & Luxol Fast Blue One of the most common stains used for nervous system tissues is the Cresyl Violet method. Cresyl Violet binds strongly to the RNA in the neuron's rough endoplasmic reticulum, since it's a basic stain. Luxol Fast Blue gives particularly good delineation of nerve tracts in the CNS, and the two are commonly used together.
Osmium Tetroxide
Osmium tetroxide (OsO4) is both a stain and a fixative. While it's primarily used these days as a fixative in electron microscopy, since it binds to lipids strongly, it's particularly well suited to reveal the details of myelin in nerves (see below). It's been used for this purpose for a very long time: the first paper reporting the use of OsO4 dates to 1865. When it reacts with the double bonds of lipids, osmium tetroxide is reduced and metallic osmium is deposited in the tissue. This gives the tissue a black coloration, and myelinated nerve fibers in particular react strongly. You'll see it below when we look at myelin.
Other Methods
There are hundreds of other staining routines, most of which involve the use of gold or silver salts. Among the most elegant of these stains are the ones developed by Camillo Golgi (1843-1926) or Santiago Ramon y Cajal (1852-1934), who shared a Nobel prize for their work in 1906. These methods are especially useful for visualizing glial elements.
 Both these men are great figures of the history of the life sciences and the study of the nervous system in particular. Golgi developed several stains that are still used today, was the discoverer of several important nervous system structures, and won the Nobel Prize for his work. Golgi's stains comprise a set of methods for nerve cells and fibers; they're characterized by fixation in an aldehyde-osmium-dichromate solution, followed by impregnation with silver salts. As you can see here, the process renders the subject as several shades of golds, browns and blacks. Neuron somata are golden and their processes black. This stain permits the definition of much detailed information about the structure of the nervous system.
Both these men are great figures of the history of the life sciences and the study of the nervous system in particular. Golgi developed several stains that are still used today, was the discoverer of several important nervous system structures, and won the Nobel Prize for his work. Golgi's stains comprise a set of methods for nerve cells and fibers; they're characterized by fixation in an aldehyde-osmium-dichromate solution, followed by impregnation with silver salts. As you can see here, the process renders the subject as several shades of golds, browns and blacks. Neuron somata are golden and their processes black. This stain permits the definition of much detailed information about the structure of the nervous system.
The Basic Unit: The Neuron
The neuron or nerve cell is the basic functional unit of the system, though they are by no means the only (or even most numerous) cell type present. Neurons come in several forms, which can be classified by their structure, their function, or both.
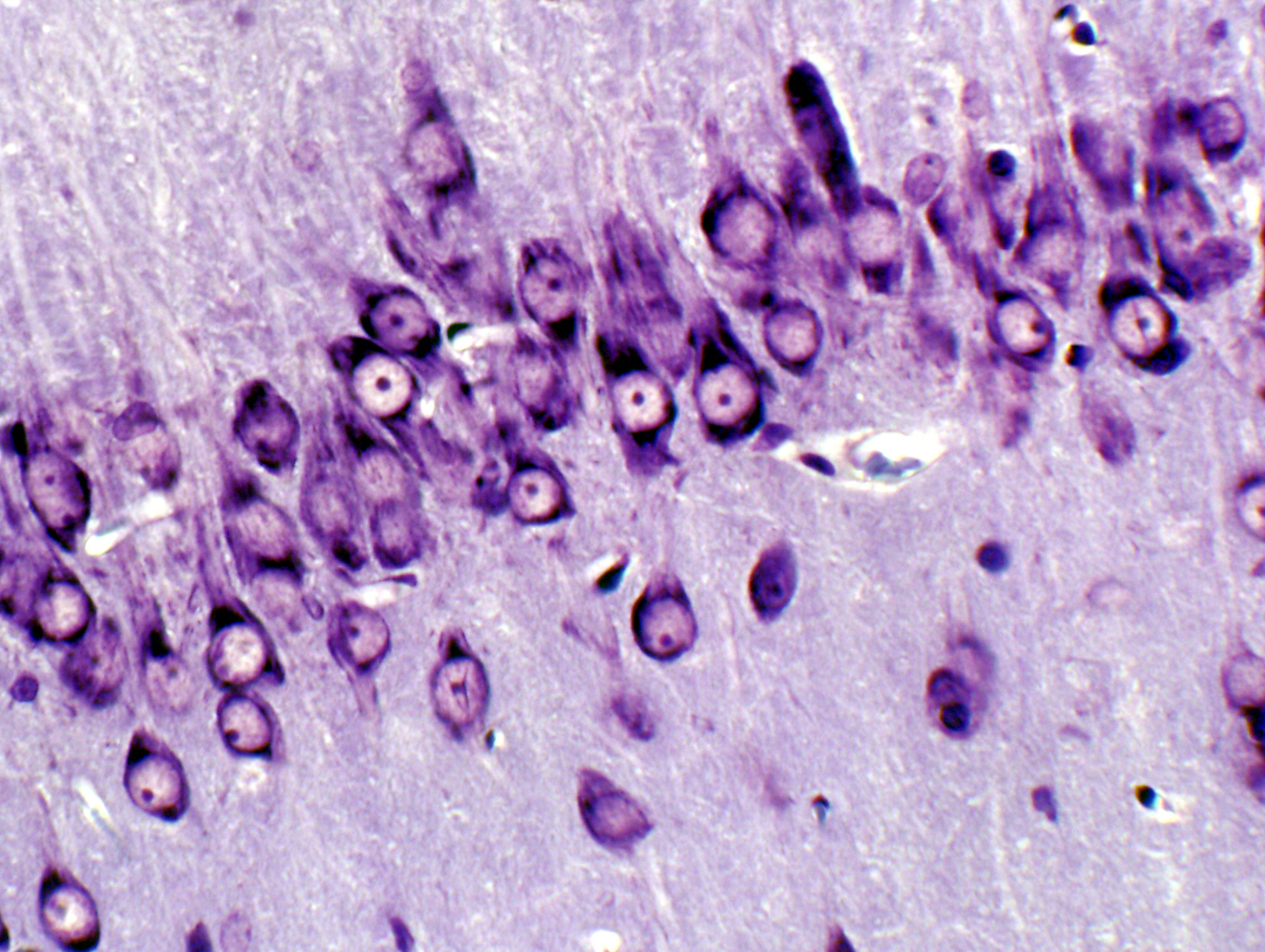
You can examine individual neurons in several of the slides in the set, especially slides 537 and 552. In these slides you'll be able to find numerous examples of neurons. These for the most part are irregularly stellate in shape. In the cerebellum and cerebrum (see below) the neurons you'll see are connector or integrator neurons, whose function is to pass information around inside the brain. The neurons of the spinal cord are classified functionally either integrator or motor neurons.
Most of the examples of neurons you'll see on slides 537 and 552 are structurally classified as multipolar. They have several projections visible in a light microscope. These projections are the axons and the dendrites of the cells; axons send a signal out of the body (soma) of the neuron, and dendrites receive input and transmit that signal to the soma. The projections of neurons (especially axons) can be very long (in some cases, several feet) so it's unusual for all of a projection to be in the same plane of section. Hence, most of what you see as the "neuron" is its soma or cell body. Most neurons are structurally classified as multipolar. Neurons that have integrator functions and/or motor functions are usually multipolar; neurons with sensory roles may be, but are more typically bipolar or "pseudo-unipolar" in structure.
In slides stained with cresyl violet, you'll make out large irregular clumps of rough endoplasmic reticulum inside the neuron somata at high magnification. Neurons are highly secretory in nature, and so they can be expected to have lots of RER and a prominent nucleolus.
Nerves
People often use the terms "neuron" and "nerve" interchangeably, but this is incorrect. A "nerve" is a grossly visible anatomic structure, and is a bundle of axonal processes from many different neurons, wrapped in a connective tissue sheath. A "neuron" is a cell, one part of which is included in a nerve.
 A telephone cable (right) makes for an apt analogy: the cable carries messages back and
forth along wires. The multi-plex, bi-directional cable is made up of wires. Each wire has its own insulation,
and each carries it's own specific separate message. There may be thousands of
wires in a cable, and they're bound together inside it by wrappings of further
insulation. Over the whole of the outside of the cable is a waterproof
covering. To continue the analogy, consider what happens when a large cable runs down a main street. Within that cable are sub-bundles, consisting of the individual lines to houses on the street, and to houses on side streets. Periodically one of the sub-bundles comes out of the main cable (the "trunk line") and enters its own street, and along that run, each single line is branched off to serve individual houses.
A telephone cable (right) makes for an apt analogy: the cable carries messages back and
forth along wires. The multi-plex, bi-directional cable is made up of wires. Each wire has its own insulation,
and each carries it's own specific separate message. There may be thousands of
wires in a cable, and they're bound together inside it by wrappings of further
insulation. Over the whole of the outside of the cable is a waterproof
covering. To continue the analogy, consider what happens when a large cable runs down a main street. Within that cable are sub-bundles, consisting of the individual lines to houses on the street, and to houses on side streets. Periodically one of the sub-bundles comes out of the main cable (the "trunk line") and enters its own street, and along that run, each single line is branched off to serve individual houses.
 So it is with nerves; the "wires" are the axonal processes
of the neurons (whose somata are elsewhere) and each "wire" has
insulation in the form of myelin and/or a Schwann cell sheath. If you compare the striking 3-dimensional scanning EM image at left to the sectioned phone cable above, you can easily appreciate the similarities, I think!
So it is with nerves; the "wires" are the axonal processes
of the neurons (whose somata are elsewhere) and each "wire" has
insulation in the form of myelin and/or a Schwann cell sheath. If you compare the striking 3-dimensional scanning EM image at left to the sectioned phone cable above, you can easily appreciate the similarities, I think!  An anatomic nerve may have thousands of axons in it; each has a specific destination. Bundles composed of these "insulated" individual axons are wrapped together with CT to form into sub-bundles of the whole cable (in larger nerves, analogous to trunk lines)
there are bundles of such bundles, also held together by CT. The whole "cable" has a tough CT sheath around it. Along the run of the main nerve, smaller branches come off, to serve smaller and smaller regions; from these arise even smaller branches, and eventually a single axon breaks out of the smallest branch to innervate its specific target. At right is an entire anatomic nerve cut in cross section: the CT that subdivides it into bundles are indicated as P (perineurium) and E (epineurium), which will be discussed at more length after we deal with the insulation on the "wires" themselves.
An anatomic nerve may have thousands of axons in it; each has a specific destination. Bundles composed of these "insulated" individual axons are wrapped together with CT to form into sub-bundles of the whole cable (in larger nerves, analogous to trunk lines)
there are bundles of such bundles, also held together by CT. The whole "cable" has a tough CT sheath around it. Along the run of the main nerve, smaller branches come off, to serve smaller and smaller regions; from these arise even smaller branches, and eventually a single axon breaks out of the smallest branch to innervate its specific target. At right is an entire anatomic nerve cut in cross section: the CT that subdivides it into bundles are indicated as P (perineurium) and E (epineurium), which will be discussed at more length after we deal with the insulation on the "wires" themselves.
Myelinated and Non-Myelinated Nerve Fibers
Axons in the periphery are never "naked." They always have some form of covering, which is the product of the activities of the Schwann cell (or, in the central nervous system, the oligodendrocyte).
Two forms of this "insulation" exist. The first type is a very elaborate multi-layered covering of plasma membrane, a thick and efficient barrier to charge leakage, easily seen with the light microscope. This is a myelin sheath. The myelin sheath is made by Schwann cells in the peripheral nervous system and by oligodendrocytes in the central nervous system. Axons with such a covering are referred to as myelinated.

 This is a medium magnification view of part of a peripheral nerve cut in cross section. It's composed of individual axons, each of which is surrounded by a myelin sheath. Myelin is very rich in lipids, so it will readily bind osmium tetroxide, with which this section has been stained.
This is a medium magnification view of part of a peripheral nerve cut in cross section. It's composed of individual axons, each of which is surrounded by a myelin sheath. Myelin is very rich in lipids, so it will readily bind osmium tetroxide, with which this section has been stained.
Note that each axon has its own myelin sheath. Just as in a telephone cable the individual wires are insulated, so too are many of the axons in a nerve fiber. In axons of equal size the rate of conduction of a signal is much higher when there's a myelin sheath. Myelination prevents leakage of membrane charge into the surrounding intercellular space. It also lessens the strain on the neuron's sodium potassium pump by restricting ion release to specific sites (see below).
In the image at right, you see nerve fibers that have been "teased" apart. Notice that the myelin sheath is discontinuous along the length of the axon. The "breaks" are minor in size and comprise only a small total of the length of the axon, but they have major functional significance. These Nodes of Ranvier (NR) will be discussed in more detail below.
An anatomic nerve can have fibers that are sensory or motor in nature, though some nerves have one kind and not the other. If a nerve runs from the tip of the toe to the spinal cord in a large animal, obviously it has to be several feet long. Within it, there are axons that are also several feet long, and every inch is covered with myelin except at these nodes. In very large animals, nerves (and hence nerve fibers, i.e., axons) two meters or more in length are normal. These fibers carry sensation to the CNS and motor commands from it to the periphery.
 The Schwann cell (or, in the CNS, the oligodendrocyte) extrudes vast quantities of plasma membrane, and wraps them around the axon, exactly the way paper towels are wound around the core of the roll. Hundreds of layers of myelin can be present: the larger the axon the thicker and more effective its myelin sheath will be. Although it's visible in the LM, The true nature of myelin can't be appreciated without the
electron microscope, and so a demonstration of myelin in the TEM is provided.
Here's a typical cross section of a myelinated nerve fiber, as seen in the light electron microscope: the nucleus of at least on Schwann cell is visible. The windings of plasma membrane around and around the axon are seen at this magnification (1000x) as a pink sheath around the axon in the center. The nucleus of the Schwann cell sits right on top of the sheath it's made.
The Schwann cell (or, in the CNS, the oligodendrocyte) extrudes vast quantities of plasma membrane, and wraps them around the axon, exactly the way paper towels are wound around the core of the roll. Hundreds of layers of myelin can be present: the larger the axon the thicker and more effective its myelin sheath will be. Although it's visible in the LM, The true nature of myelin can't be appreciated without the
electron microscope, and so a demonstration of myelin in the TEM is provided.
Here's a typical cross section of a myelinated nerve fiber, as seen in the light electron microscope: the nucleus of at least on Schwann cell is visible. The windings of plasma membrane around and around the axon are seen at this magnification (1000x) as a pink sheath around the axon in the center. The nucleus of the Schwann cell sits right on top of the sheath it's made.
 The spaces in between layers of plasma membrane contain no cytoplasm. Compare the H&E stained image above with the transmission electron micrograph shown at right. This TEM preparation is at about 25,000 diameters and shows details of the sheath's nature and its relationship to the bulk of the Schwann cell.
The spaces in between layers of plasma membrane contain no cytoplasm. Compare the H&E stained image above with the transmission electron micrograph shown at right. This TEM preparation is at about 25,000 diameters and shows details of the sheath's nature and its relationship to the bulk of the Schwann cell.
Again, you're seeing a Schwann cell perched on an internodal segment of the myelin sheath, cut in cross section; but at this magnification it's possible to see the layering of the sheath and the relationship of the nucleus to the wrapping. The cytoplasm of the Schwann cell (which contains all the usual organelles you'd expect) surrounds the nucleus.
The axonal cytoplasm (axoplasm) is pretty bland looking. The large oval object in it is a mitochondrion. Mitochondria are numerous in the axon, providing the power to run the sodium potassium pump that keeps the transmembrane potential at normal levels. The small circular profiles in the axoplasm are microtubules. These maintain normal axonal shape, and guide neurotransmitter molecules synthesized in the soma to their destination in the terminus of the axon. The very high lipid content of the membrane wrapping is what gives the sheath its intense staining reaction with osmium.
Unmyelinated Axons
 Not all axons of peripheral nerves are myelinated. But that doesn't mean they're uncovered; neurons are never left naked. In the case of non-myelinated
axons the Schwann cells don't wrap themselves around the process
hundreds of times; they simply envelop it with one fold of their
plasma membrane instead of many. Unmyelinated axons are usually fairly small,
and the difference between myelinated and unmyelinated ones is easily
appreciated in the accompanying demonstration of an unmyelinated axon in the
TEM. Two myelinated axons, both much larger, are in the image for comparison. This picture was taken at about 50,000x magnification.
Not all axons of peripheral nerves are myelinated. But that doesn't mean they're uncovered; neurons are never left naked. In the case of non-myelinated
axons the Schwann cells don't wrap themselves around the process
hundreds of times; they simply envelop it with one fold of their
plasma membrane instead of many. Unmyelinated axons are usually fairly small,
and the difference between myelinated and unmyelinated ones is easily
appreciated in the accompanying demonstration of an unmyelinated axon in the
TEM. Two myelinated axons, both much larger, are in the image for comparison. This picture was taken at about 50,000x magnification.
What's myelin for? Why elaborate such a complex and energetically expensive material? Myelination increases conduction velocity of nerve processes. Remember that the conduction along an axon is electrical in nature, and that like any such "wire" if there is a "short circuit" function is interrupted. The insulating myelin prevents charge leakage, signal loss, and interference with normal function.
Nodes of Ranvier

 As has been mentioned, the myelin sheath has breaks in it, the nodes of Ranvier (NR). The nodes are easily visible in a teased preparation, but somewhat harder to spot in ordinary sections. However, there are so many in an anatomic nerve that almost any longitudinal section, such as this one, will show a few.
As has been mentioned, the myelin sheath has breaks in it, the nodes of Ranvier (NR). The nodes are easily visible in a teased preparation, but somewhat harder to spot in ordinary sections. However, there are so many in an anatomic nerve that almost any longitudinal section, such as this one, will show a few.
In the peripheral nervous system internodal segments of myelin (i.e., the product of one Schwann cell's activity) cover a few microns to a few millimeters; then there's a node, before the next segment begins. In other words, the node is located between one Schwann cell and the next. In the image below and at right, which is stained with Mallory's connective tissue stain, you can make out the relationship of the myelin sheath at the node and the axon: the axon runs continuously through the node from one internodal segment to the next, for its full distance.
Turn to slide 540. This is a cross-section of peripheral nerve. The nerve (and of course also the axons inside it) are cut in cross section. In most cases a myelin sheath is visible around each axon. If you find one wherein the Schwann cell nucleus can be seen closely pressed against the axon, look at it under oil immersion. You will clearly see the relationship of the cell, the axon, and the myelin, which in H&E preparations appears as a clear band around an apparently "empty" axon.
 Now let's go back to the telephone cable. Several gradations of CT envelopments are also visible. The endoneurium surrounds individual axons; if the myelin sheath is insulation, the endoneurium is the fabric cover of the "wire." The endoneurium is confluent with the perineurium, CT that ties bundles of axons together; and the perineurium eventually grades into the epineurium, the CT that binds the whole
"cable" into a unit.
Now let's go back to the telephone cable. Several gradations of CT envelopments are also visible. The endoneurium surrounds individual axons; if the myelin sheath is insulation, the endoneurium is the fabric cover of the "wire." The endoneurium is confluent with the perineurium, CT that ties bundles of axons together; and the perineurium eventually grades into the epineurium, the CT that binds the whole
"cable" into a unit.
At left is a cross section, stained with a CT routine to highlight the relationship between the axons and the various levels of connective tissue around them. Each axon is invested with a wispy covering of delicate reticular CT, the endoneurium. This in turn is "tied" to the endoneurium of adjacent axons, and becomes at some point the perineurium. Perineurium has several hierarchical levels in turn, and eventually it will merge with the epineurium surrounding the outside of the anatomic nerve. This relationship can be studied on slide 8; the image above is made from a demonstration slide cut from the same block.
Slide 270 is a longitudinal section, approximately on the midline, of an entire rat brain. It's stained with a combination of Luxol fast blue and the PAS stain. Since almost all of it present on this slide, and it's best examined grossly first to orient yourself. If you then examine the slide under low power (4x or lower) you'll be able to make out the general layout. Orient it on your stage so that as you view it the cerebellum is to the right.bThe most cranial end is the olfactory lobe, and the cerebral hemispheres can easily be made out as well. Caudally, the most prominent structure is the cerebellum, which somewhat resembles a tree or leaf. (In fact, one of the older terms for this structure was arbor vitae, Latin for "tree of life," reflecting the idea that without it one would die, both spiritually and physically. They were right, too.)
These two terms always cause some confusion, but they're so deeply entrenched in the literature nothing will ever dislodge them; so I suppose we should discuss them here. Strictly speaking, the terms are valid only in the context of gross anatomy. If you slice open a brain or a spinal cord, the freshly cut surfaces have some areas which are a glistening white color, and some a sort of duller gray shade. The term white matter is used to describe the former; it consists of areas that are primarily composed of nerve fiber tracts. The gray matter represents areas in which the number of neurons is high. Since neurons tend to accumulate lipofuscin and in some cases, melanin, they stand out against the white "background" of fiber tracts.

The distinction between gray and white matter isn't so clear-cut in the microscope as it is on the dissecting table. Even under low magnification you'll see fiber tracts running through areas of "gray matter." Those neurons have inputs and outputs, just as all neurons do, and signals are carried to and from them by axons and dendrites. The microscope brings you so "close" to the preparation that the white versus gray distinction is lost. However: we can define the terms in histological terms as follows:
In the central nervous system, "gray matter" is the term applied to neuron cell bodies in the aggregate, usually those clustered together in the nuclei of the brain and spinal cord. "White matter" is everything else.
Neuron cell bodies in the gray matter can easily be seen in the outer portions of the olfactory lobe and of the cerebrum on slide 270. This arrangement of gray matter on the outside is the reverse of the way things are arranged in the spinal cord (see below). In the cord the gray matter is in the center and the white matter around it. There's a region of transition at the caudal end of the brain, visible on slide 270. These regional differences in architecture come about due to migration of cells during the embryological development of the neural tube into its final form.
The cerebellum can be examined on slide 270 and also in somewhat more detail in slide 543, which has been stained with a silver impregnation method. Examined grossly, it's "leaf-like" in organization. Under moderate magnification, it can be seen that this appearance comes from the folding and convolution of its cortex into folia, or "leaves"—as in the "leaves" of a book.

 This is a low magnification image of most of the cerebellum.
This is a low magnification image of most of the cerebellum.
The cerebellum has a cortex and a medulla, and what you see in this image is mostly cortex. It's folded like a piece of cloth, so that if you could follow the outermost surface you would dip down between the leaf-like folds (the foliae, F) and back up again. The medulla consists of nerve fibers (NF) leading out of the cortex.
The cerebellar cortex has three layers, only two of which can be made out at this magnification. The outermost molecular layer, the pink band along the surface, is a vast array of nerve fiber tracts. These interact with cells in the next layer, which isn't visible here. The innermost layer of the cerebellum, the granule cell layer is seen here as a darker-staining region, stippled with nuclei.
Molecular and Purkinje Cell Layers of the Cerebellum
The outermost layer of the cerebellar cortex is the molecular layer, a region relatively free of neuronal somata. The molecular layer is composed of the projected axons of the cells of the granule cell layer (see below) and the immensely ramified dendritic trees of the Purkinje cells (see below).

Immediately deep to the molecular layer is one composed of the very large Purkinje cells, discovered by and named after Johannes G.J. Purkinje (1787-1869), a great Bohemian (i.e., Czech) physiologist and anatomist. Purkinje cells are big, even by nervous system standards. They average 150-200 µm in diameter. In a two-dimensional section you see them lined up neatly into a single row, but realize that this is a spurious appearance arising from the two-dimensional nature of microscopic slide sections. Purkinje cells really form a layer. Thousands of them are aligned into ranks and rows forming a grid-like construct between the molecular layer and the underlying granule cells.

These images only give the faintest hint of the fantastic array of Purkinje cell's dendrites. The base of the "tree" where it's rooted to the cell soma is visible. Each "branch" of the tree is ramified into tens of thousands of smaller branches, forming a colossal fan-like array that extends up into the molecular layer. Each Purkinje cell's extensive dendritic array extends upwards into the molecular layer, oriented at right angles to the direction in which the fibers of the molecular layer run. There are hundreds of thousands—even millions—of points of contact between the molecular layer axons and the dendrites of each Purkinje cell.
The Purkinje cells are the principal integrating neurons of the cerebellum, the classic example of an integrator. From each a large axon descends into the deeper portions of the cerebellum. Its function is to receive input from various sources, make a "decision" to fire or not, and to send a single integrated on/off signal from its own axon reflecting that decision. The arrangement of the Purkinje cell's dendrites and their relationship to the nerve fibers in the molecular layer are the important factors in making the system work.
The vast complexity of the synaptic contacts between the Purkinje cells and the fibers of the molecular layer provides for a rapid and simultaneous processing of signals. In Computerspeak, we'd described this as a "massively parallel processing system" in which huge amounts of information can be handled in a very short period. The cerebellum is constantly being fed inputs from proprioceptor organs, and visual cues, and sound information, etc. It needs this capability of almost instantaneous signal input and output to function properly.
The molecular layer itself consists of millions of fibers, which pass through the serried ranks of Purkinje cell dendrites like telephone wires running through the trees of an orchard. At every point where a "wire" contacts a "branch" there is a synaptic connection: a single Purkinje cell may easily have tens of thousands of such inputs. Some of these synapses are excitatory, others inhibitory, and the possible combinations on a given Purkinje cell are in the trillions. Multiply that by the tens of thousands of Purkinje cells, and you can readily see that the possibilities of integrating input and output are nearly infinite.
Equally important, the time required for the system to process millions of inputs and make a coordinated set of outputs take place is very short. The final output of any given Purkinje cell is via a single axon, but all the Purkinje cells are doing this simultaneously, taking sensory information from all parts of the body and sending output to the appropriate parts of the CNS that control the effectors.
The Purkinje cell axons pass through the granule cell layer and form the fiber tracts of the cerebellar medulla. Information is routed to other parts of the CNS, where more decisions are made and final motor output is determined. This colossal parallel processing capability allows almost instantaneous response. The remarkable acrobatic ability of many arboreal animals such as squirrels (and the predatory species who hunt them) is due to a highly developed cerebellum. If a cat happens to fall off something (say, the top of a china cabinet) she will always land on her feet. The cerebellum processes positional information, muscle tonus, acceleration data, visual clues, and hundreds of other inputs in milliseconds, so that the cat can twist in mid-fall to land properly. You don't have to take my word for it: find a cat, throw her up in the air, and see what happens. (Better not stand underneath where she is likely to land, because her cerebellum will also cause her to unsheathe her claws.)
Cerebellar control of movement isn't really "automatic" in the same sense that breathing is. Breathing is truly automatic and a newborn baby can breathe properly. Breathing is so fundamental that it's controlled by neural mechanisms in the most ancient parts of the brain, and even individuals who've lost all cerebellar function are able to breathe properly. But animals have to learn coordination, "conditioning" the neural circuits of the cerebellum with the appropriate paths for sensation and response. It takes time to learn how to walk and chew your cud at the same time, so to speak.
The time varies considerably among species. Some animals (especially herd animals) are born with a fully-developed cerebellum, and the capability for coordinated movement is there almost from the moment they hit the ground. Calves and foals have to keep up with the herd or they'll die. The helplessness of kittens and puppies, who require weeks to learn to walk and run, is due to the fact that in those animals the development of the cerebellum isn't completed until a few weeks after birth. Typically, in such species the parental level of care-giving is quite prolonged. In humans the process of learning to walk and run takes a year or more.
However, once the techniques of postural and movement control mastered, barring injury or disease, they become as "automatic" as breathing, for all practical purposes. If the cerebellum doesn't develop fully, an animal may never develop proper coordination. A variety of conditions may result in cerebellar hypoplasia. An animal with an improperly formed or under-developed cerebellum will be clumsy and uncoordinated. Sometimes they may actually fall face-first into their food bowls.
Immediately deep to the Purkinje cell layer are the somata of the cells whose axons form the bulk of the molecular layer. These are small neurons called granule cells, forming a compact and easily distinguished region. In embryonic development these cells actually arise at the outer surface of the neural tube, but they migrate past the developing Purkinje cells, leaving a long axon behind them as they go.
Deep to the granule cell layer is the cerebellar medulla, made up of the axonal processes of the Purkinje cells. These pass through the granule cell layer and become bundled together into fiber tracts.
Notice that all three of the layers of the cortex, and the underlying layer of fibers, is present in each of the foliae. You can trace the contours of a layer from one end of the cerebellum to the other without ever leaving that layer. These structures and relationships can be seen also in slide 544.
At first glance, you might think 552 was another slide of cerebellum, but compare it to the slides of cerebellum and you'll see the differences. A good many neurons are visible in this slide, but the organization is much different. These neurons are layered, but all of the layers are on the outer portion of the section; there is white matter underneath, just as in slides 270 A&B. Of course, there are no Purkinje cells.
The convolutions and folds that give the cerebrum a superficial resemblance to the cerebellum are the gyri. It's easy to tell the two apart, however, because of the unique layering of the cerebellum and the very large Purkinje cells found there.
 The central nervous system of vertebrates is "hollow." It begins as a
closed tube in the early embryo, and even after its final development it retains a
lumen, the remains of that of the primitive neural tube. This hollow region is filled with cerebrospinal
fluid, produced by a special secretory structure that's present in various locations inside that
lumen. On slide 270 you should be able to make out at least two places where this
structure, the choroid plexus, is visible inside the brain. A plexus is present in both lateral ventricles (the hollow spaces of the cerebral hemispheres, not visible on this slide). On slide 270 you can make out a plexus at the cranial end in the space below the
cerebrum (the third ventricle). There's one located more caudally, below
the cerebellum (in the roof of the fourth ventricle).
The central nervous system of vertebrates is "hollow." It begins as a
closed tube in the early embryo, and even after its final development it retains a
lumen, the remains of that of the primitive neural tube. This hollow region is filled with cerebrospinal
fluid, produced by a special secretory structure that's present in various locations inside that
lumen. On slide 270 you should be able to make out at least two places where this
structure, the choroid plexus, is visible inside the brain. A plexus is present in both lateral ventricles (the hollow spaces of the cerebral hemispheres, not visible on this slide). On slide 270 you can make out a plexus at the cranial end in the space below the
cerebrum (the third ventricle). There's one located more caudally, below
the cerebellum (in the roof of the fourth ventricle).

Each plexus consists of a tortuous loop or tuft of capillaries, overlain by a neatly cuboidal epithelial covering. The plexi protrude into the fluid-filled lumen of the ventricle. In the electron microscope, the cells covering the blood vessel have a sort of brush border consisting of irregular microvilli. This is in keeping with their function: producing the cerebrospinal fluid that fills the ventricles of the brain and the lumen of the spinal cord.
The choroid plexus capillaries are covered by a special cuboidal epithelium. The cells of this epithelium are sealed together by occluding junctions, to maintain the integrity of the blood-brain barrier.
Using materials brought in the circulation, the cells of the choroid plexus synthesize the components of the cerebrospinal fluid and secrete the final product into the lumen of the ventricles. The occluding junctions prevent unregulated movement of material from the blood compartment into the CSF-filled spaces. Unless it's mediated by the cells of the plexus, transfer doesn't take place.

Turn now to slides 537 and 538. The first is a cross section of spinal cord stained with cresyl violet. The second is silver-stained. Slide 537 bears both a cross section and a longitudinal section.
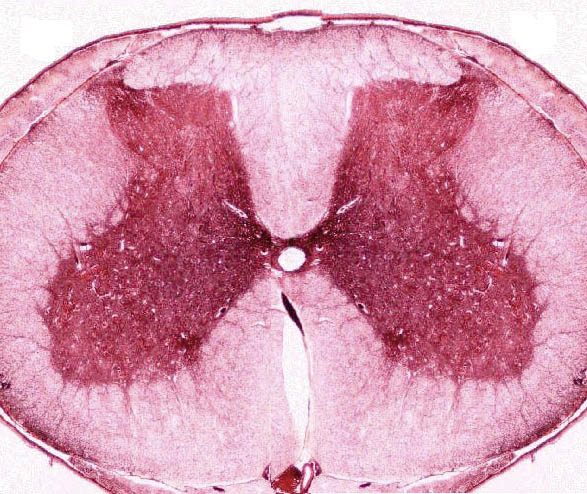
The cord is arranged with "white matter" on the outside and the "gray matter" in the center. In the cross section, you'll easily see that the gray matter takes an H shape, with a small channel, the central canal in the middle. The central canal, like the brain ventricles, is filled with cerebrospinal fluid and lined with ciliated cells that circulate the fluid.
The architecture of the neurons and their axons is beautifully defined by the silver staining method, and there's probably no other slide in this set which so clearly delineates the relationships between somata and the processes to which they give rise. Fiber tracts are especially clear in this slide.
The spinal cord is essentially a large communications cable, and most of it consists of axonal projections running up and down to and from the brain.
The central canal is the remaining lumen of the hollow embryonic neural tube. The cells lining it are the ependymal cells, whose function is the circulation of cerebrospinal fluid. Ependymal cells also line the ventricles of the brain. They're ciliated and the movement of the cilia circulates CSF throughout the hollow spaces of brain and cord. Ependymal cells are tall and usually described as "epithelioid" in nature. "Epithelioid" is Anatomyspeak for, "It looks like epithelium but it doesn't have a basement membrane, so it isn't." Ependymal cells aren't epithelium but they do a lot of the same things true epithelial cells do and they're built the same way, so they're "epithelial-like," or epithelioid.
The fiber tracts of the white matter are pretty obvious; keep in mind that in a cross section, you are seeing more axons cut transversely than longitudinally. Sometimes you'll be able to make out the myelin sheaths on these axons.
Within the gray matter's H or "butterfly" shape, the ventral horns of the gray matter are larger than the dorsal horns. In the ventral horn lie the large somata of autonomic motor neurons. These cells receive input from integrator neurons in the cord, and send commands to contract to skeletal muscle in the periphery.
The dorsal horn of the cord's gray matter contains neuron somata, also; but they aren't motor in function. These are the somata of the connector neurons. They receive input from sensory fibers entering the cord through the dorsal root of the spinal nerve.
A complete somatic reflex arc consists of 1) a sensory structure; 2) a sensory neuron; 3) an integrator neuron whose soma is in the gray matter of the cord's dorsal horn; 4) a motor neuron whose cell body is in the ventral horn of the cord's gray matter; and 5) an effector organ, in the form of skeletal muscle.
Autonomic reflex arcs also exist. These have the same sort of sensory input (a sensory structure and its associated sensory nerve), but the motor output differs. There are two neurons in the motor chain, one of which is located in the spinal cord, and the second of which is in an autonomic ganglion of the peripheral nervous system.
The meninges, the specialized covering of the CNS, cover the cord as well as the brain. The meninges on the spinal cord are visible on slide 539, which has been stained with both H&E and the PAS/Alcian Blue reactions. The same three meningeal coverings are present around the brain, but they're easier to define in a section of spinal cord.

The outermost meningus, the dura mater (DM) is a tough, dense CT envelope. The name is Latin, for "tough mother," (!) which seems appropriate. Grossly it looks like tough wet paper, and in a slide stained for connective tissue, it's a dense band, as you see it on slide 539.
Deep to the dura is the arachnoid meningus (from the Greek arachne, a spider or cobweb), a delicate irregular CT whose interstices are filled with cerebrospinal fluid, and through which blood vessels are routed. In this image it's labeled Ar.
 The third
meningus is the pia mater (from Latin again, "soft mother") a very thin and delicate CT closely adherent to
the surface of the brain and spinal cord. In the image above it can barely be made out as a delicate blue edge on the white matter (WM) of the ventral surface of the cord. The pia mater follows every curve and
convolution of the surface. This can be seen at right.
The third
meningus is the pia mater (from Latin again, "soft mother") a very thin and delicate CT closely adherent to
the surface of the brain and spinal cord. In the image above it can barely be made out as a delicate blue edge on the white matter (WM) of the ventral surface of the cord. The pia mater follows every curve and
convolution of the surface. This can be seen at right.
On the inside surface of the pia mater, where it faces the actual brain tissue space, foot processes of astrocytes (see below) form a limiting structure, the glia limitans, which is part of the blood-brain barrier. Since the meninges are CT they have blood vessels in them and neurons have to be sheltered from direct contact with substances in the blood.
Notice that on slide 539, there's also a large dorsal root ganglion near the cord. We'll come back to it again (see below).
To continue the telephone analogy, so far we've seen the "wiring" (the nerves) and the "central exchange" (the brain) and the "main trunk line" (the spinal cord). So let's look at the intermediate structures that route the signals around in subdivisions of the body.
Nerves, like telephone cables, occasionally run into "junction boxes" where the signals they carry are routed to the appropriate places. In the peripheral nervous system the "junction boxes" are ganglia (singular form is "ganglion"). A "ganglion" by definition is any collection of neuron cell bodies outside the central nervous system. Although aggregations of neuron cell bodies exist in the central nervous system (roughly analogous collections called "nuclei") these aren't ganglia. But all neuron cell bodies outside the brain and spinal cord are in ganglia.
There are two principal types, distinguished mainly by their function and sometimes by location. One type is sensory, the other motor.
The first type of ganglion is the craniospinal ganglion. As the name implies, these are associated with the spinal cord and the cranial nerves
There's a craniospinal ganglion on slide 539. It's near one side of the spinal cord. This particular example can also be correctly termed a "dorsal root" ganglion, so called because it's on the dorsal root of a spinal nerve. This type of sensory ganglion is also found on cranial nerves, hence the term "craniospinal" is appropriate for either location. "Dorsal root ganglion" is appropriate only for the ones on the spinal nerves.
Craniospinal ganglia are sensory in function. In the case of dorsal root ganglion, the cell bodies inside it are those of "pseudo-unipolar" neurons. This structural class of neurons is always associated with sensory functions. The name "pseudo-unipolar" refers to the spurious appearance in light microscopy of a single process arising from the soma. In fact, these neurons have two processes, but close to the soma they're fused so they appear to be only one.
One process—the afferent fiber—comes into the soma from a sensory structure in the periphery. This can be any kind of sensory structure: a touch receptor, cold receptor, pain fiber, pressure sensor, etc. The sole function of this myelinated afferent fiber is to bring the information from the periphery to the ganglion, and more exactly, into the soma of its neuron.
The second fiber—the efferent fiber—leaves the soma in the ganglion and carries the information into the central nervous system for further action. It, too, is a myelinated fiber. In most cases it exits the anatomic ganglion and runs back into the CNS via the dorsal root of the spinal nerve. Its sole function is to relay the information from the periphery to the "decision-making" regions of the CNS.
Neuron somata in craniospinal ganglia are usually very large: 100 to 150 microns in diameter. They have correspondingly large round nuclei, that are centrally located in the cell. Each soma has associated with it about 4 to 6 smaller nuclei, or satellite cells. These are supporting cells analogous in some ways to the astrocytes of the CNS (see below). They're called satellite cells.
 This is a low power view of a sensory ganglion. This particular example is a dorsal root ganglion from one of the spinal nerves. Sensory ganglia of similar structure are also found on cranial nerves.
This is a low power view of a sensory ganglion. This particular example is a dorsal root ganglion from one of the spinal nerves. Sensory ganglia of similar structure are also found on cranial nerves.
A ganglion is a discrete structure, with a well-defined CT outer capsule (C). Inside the ganglion are neuron cell bodies. Neurons have to be wired into the system, of course, so nerve fibers (NF) are always present. In the case of craniospinal/sensory ganglia, these fibers are the ones bringing information in from the peripheral sensory sites, and those carrying that information onward to the central nervous system for further processing.
 At higher power view you can make out some more details. The large neurons in it are round in shape (usually only the soma is visible) with centrally located nuclei. Nucleoli are usually present. Like any neurons, these have a retinue of glial cells to dance in attendance on them and cater to their whims. In this case the glial element is the satellite cell. These are for all intents and purposes the peripheral nervous system's equivalent of the astrocyte, with many of the same functions. In a craniospinal ganglion four or so satellite cells are associated with each neuron, their nuclei are arranged around the outside of that neuron's "personal space." The satellite cells form a sort of individual "glia limitans" for each neuron, just as the astrocytes do for the neurons of the brain.
At higher power view you can make out some more details. The large neurons in it are round in shape (usually only the soma is visible) with centrally located nuclei. Nucleoli are usually present. Like any neurons, these have a retinue of glial cells to dance in attendance on them and cater to their whims. In this case the glial element is the satellite cell. These are for all intents and purposes the peripheral nervous system's equivalent of the astrocyte, with many of the same functions. In a craniospinal ganglion four or so satellite cells are associated with each neuron, their nuclei are arranged around the outside of that neuron's "personal space." The satellite cells form a sort of individual "glia limitans" for each neuron, just as the astrocytes do for the neurons of the brain.
 Unlike the neurons characteristic of the CNS, the ones in sensory ganglia aren't stellate in shape. They're pseudo-unipolar neurons. "Pseudo-unipolar" implies that these neurons appear to have only one process, and that the appearance is spurious. Examination with an electron microscope or special stains can demonstrate that there are two processes (an afferent and an efferent process) which are so close together as to give the impression of only one, at least within the substance of the ganglion. One process is used to bring signals to the soma, the other to take them out; hence information flows through this type of neuron in only one direction: from periphery to CNS. The afferent fiber can be several meters long. The efferent fiber is only long enough to connect the soma to its input site in the CNS. There it forms a synapse with an interconnector neuron, usually in the dorsal horn of the grey matter of the spinal cord.
Unlike the neurons characteristic of the CNS, the ones in sensory ganglia aren't stellate in shape. They're pseudo-unipolar neurons. "Pseudo-unipolar" implies that these neurons appear to have only one process, and that the appearance is spurious. Examination with an electron microscope or special stains can demonstrate that there are two processes (an afferent and an efferent process) which are so close together as to give the impression of only one, at least within the substance of the ganglion. One process is used to bring signals to the soma, the other to take them out; hence information flows through this type of neuron in only one direction: from periphery to CNS. The afferent fiber can be several meters long. The efferent fiber is only long enough to connect the soma to its input site in the CNS. There it forms a synapse with an interconnector neuron, usually in the dorsal horn of the grey matter of the spinal cord.

The second type of ganglion is the autonomic ganglion, seen in slide 550. The typical place to find such ganglia is strung out along the sympathetic trunk. Autonomic ganglia are involved in the regulation of automatic functions like perspiration and blood flow. They're functionally motor ganglia, not sensory like the craniospinal type. This is an autonomic ganglion, presented here at the same magnification as the craniospinal ganglion you saw earlier. If you compare the two you will see this is about half the size of the sensory ganglion. This is part of the sympathetic trunk, the long string of ganglia lined up along the vertebral column from the base of the skull to the coccyx and connected to the spinal nerves.
 Autonomic ganglia contain the somata of the second neuron in
the autonomic motor chain (see the discussion of the autonomic reflex arc). The neurons in an autonomic ganglion receive input from the CNS, process it, and send the signal to some effector organ other than skeletal muscle. (Skeletal muscle is innervated by fibers from the ventral horn of the gray matter in the spinal cord: see Exercise 10.) Typical effector organs for autonomic ganglia are sweat glands, salivary glands, smooth muscle, etc. The neuron somata in autonomic ganglia are smaller than those
of a sensory ganglion, and their nuclei are more eccentric. There are fewer
satellite cells, only 2 or so per soma. At higher power—and again, this is the same magnification used for
the comparable view of the sensory ganglion—you will note that the
cells of this ganglion are much smaller than those of the sensory
type, and that their nuclei tend to be eccentric in the somata. They
have far fewer satellite cells, perhaps 2 or so at most.
Autonomic ganglia contain the somata of the second neuron in
the autonomic motor chain (see the discussion of the autonomic reflex arc). The neurons in an autonomic ganglion receive input from the CNS, process it, and send the signal to some effector organ other than skeletal muscle. (Skeletal muscle is innervated by fibers from the ventral horn of the gray matter in the spinal cord: see Exercise 10.) Typical effector organs for autonomic ganglia are sweat glands, salivary glands, smooth muscle, etc. The neuron somata in autonomic ganglia are smaller than those
of a sensory ganglion, and their nuclei are more eccentric. There are fewer
satellite cells, only 2 or so per soma. At higher power—and again, this is the same magnification used for
the comparable view of the sensory ganglion—you will note that the
cells of this ganglion are much smaller than those of the sensory
type, and that their nuclei tend to be eccentric in the somata. They
have far fewer satellite cells, perhaps 2 or so at most.
 Although the example provided is one that is from the sympathetic trunk ranged
alongside the spine, anatomically there are several places you can find them.
Autonomic ganglia may actually be embedded in the substance of a visceral
organ, in which case the term "terminal ganglion" is applied to them. These have very short axons that leave the ganglion to innervate their target
effector organ. One such is visible in most slides of the tongue: there are many salivary glands embedded in the tongue's mass, and these terminal ganglia serve those effectors (See Exercise 17). That's the one shown at left. There's a mass of salivary gland tissue at top left in this field; five neurons and some myelinated nerve fibers that bring signals in and out are present on the right. (All ganglia have nerves running into and out of them, so as you examine these sections, look for myelinated fibers. You
can probably find a node of Ranvier if you find an area cut
longitudinally.) Note that the nuclei of these neurons are eccentric and that there's only one or two satellite cells present: a typical "terminal" autonomic ganglion, here.
Although the example provided is one that is from the sympathetic trunk ranged
alongside the spine, anatomically there are several places you can find them.
Autonomic ganglia may actually be embedded in the substance of a visceral
organ, in which case the term "terminal ganglion" is applied to them. These have very short axons that leave the ganglion to innervate their target
effector organ. One such is visible in most slides of the tongue: there are many salivary glands embedded in the tongue's mass, and these terminal ganglia serve those effectors (See Exercise 17). That's the one shown at left. There's a mass of salivary gland tissue at top left in this field; five neurons and some myelinated nerve fibers that bring signals in and out are present on the right. (All ganglia have nerves running into and out of them, so as you examine these sections, look for myelinated fibers. You
can probably find a node of Ranvier if you find an area cut
longitudinally.) Note that the nuclei of these neurons are eccentric and that there's only one or two satellite cells present: a typical "terminal" autonomic ganglion, here.
Some autonomic ganglia are so large and so consistent in their location that they've got their own anatomic names (e.g., the cervicothoracic ganglion) and you've probably seen these in Gross Anatomy. These named structures are "collateral" ganglia. But internally, all autonomic ganglia, regardless of location, share the common structural patterns described above.
The neurons are only one of the two major cell populations of the nervous system, and they're by no means the most numerous class. The other population—in the aggregate some 10 to 50 times larger, in total numbers—is the neuroglia. These are the supportive cells of the system, and there are several types.
It's important to realize that neurons never are "naked" and thus directly exposed to the potentially hostile Real World. Neurons, like small children, can't safely be left unguarded, because they are very incompetent cells in some ways. Neurons are so highly specialized for their primary functions of impulse conduction and secretion that they're pretty much incapable of doing anything else. They're extremely vulnerable to fluctuations and changes in their environment. Of necessity, glial cells of various types must always be present to cosset and protect them, to tell them what good little neurons they are, and generally to keep their spirits up. "A happy neuron is a functional neuron," as the old saying goes. Glial cells are cheerleaders, nursemaids, nannies, and cleaner-upper cells whose duties are to tend a neuron and keep it fat, dumb, and happy. Most neurophysiology and histology texts pretty much ignore the glial cells, but the principle that "If it weren't needed it wouldn't be there," applies to these as well as any other cell population. Even though complete knowledge of glial cell functions is lacking, what they do is undoubtedly important to normal function.
Astrocytes ("star cells," so called from their shape) are the most numerous glial component of the CNS, and there are many more astrocytes than neurons. Classically a distinction has long been made between "protoplasmic" and "fibrous" astrocytes based on their morphology; but it now seems that these are really two manifestations of the same cell type. For our purposes we will simply identify "astrocytes" as a single population.
 Slide 508 is stained with Golgi's gold method. The very large black blobs in
this slide are neurons; near them you will see some smaller and stellate cells,
with numerous branched projections. These are the astrocytes. Slide 509 shows
astrocytes stained with a different method. Again, look for them near neurons. In this image there are two neurons and three astrocytes. The neurons are the very large black blobs, and the astrocytes are the stellate smaller blobs "attending" them. One astrocyte is visible next to the large neuron at upper left; and two more are associated with the neuron at lower right.
Slide 508 is stained with Golgi's gold method. The very large black blobs in
this slide are neurons; near them you will see some smaller and stellate cells,
with numerous branched projections. These are the astrocytes. Slide 509 shows
astrocytes stained with a different method. Again, look for them near neurons. In this image there are two neurons and three astrocytes. The neurons are the very large black blobs, and the astrocytes are the stellate smaller blobs "attending" them. One astrocyte is visible next to the large neuron at upper left; and two more are associated with the neuron at lower right.
 Astrocytes have extensive "foot processes" and that's what creates the stellate look. These processes are pressed up against the soma of a neuron to cover every square micron of surface where there is not actually a direct synapse. The neuron is thus, in effect, physically isolated into its own little compartment.
Astrocytes have extensive "foot processes" and that's what creates the stellate look. These processes are pressed up against the soma of a neuron to cover every square micron of surface where there is not actually a direct synapse. The neuron is thus, in effect, physically isolated into its own little compartment.
Astrocytes are believed to engage in (among other things) what's called spatial buffering of the immediate environment of neurons and synapses. When a neuron's plasma membrane depolarizes, ions are released into the space around it. Potassium ions flow out and sodium ions flow in. The localized changes in ionic strength of potassium and sodium associated with neuron depolarization have to be rapidly corrected. If the local concentration of potassium in the intercellular space becomes too great, the rapid depolarization of that segment of membrane becomes more difficult. Enter the astrocyte; it can take potassium (and other ions) into its foot processes, and transport those ions to another location. It moves them through its own soma and "dumps" them where they won't interfere with the astrocyte's neuron, hence "spatial" buffering. The ions are moved from place to place.
 Another vital role of the astrocyte is to form part of the blood-brain barrier, by butting their foot processes up against not only the blood vessels but the inner surface of the pia mater. Blood vessels carry many materials that may be harmful to neurons, and the
astrocytes, by establishing a physical presence along all such vessels and
interposing themselves in a position to "filter" incoming materials,
can stabilize the environment easily. This barrier—on blood vessels and the pia mater—is collectively the glia limitans.
Another vital role of the astrocyte is to form part of the blood-brain barrier, by butting their foot processes up against not only the blood vessels but the inner surface of the pia mater. Blood vessels carry many materials that may be harmful to neurons, and the
astrocytes, by establishing a physical presence along all such vessels and
interposing themselves in a position to "filter" incoming materials,
can stabilize the environment easily. This barrier—on blood vessels and the pia mater—is collectively the glia limitans.
Slide 509 is a particularly nice example of how this system works. You will be able to see on this slide that astrocytes have their foot processes lined up along blood vessels that run through the brain. Astrocytes have similar processes that butt up against, surround, and protect neuronal somata. There isn't any "open" space between neurons: the astrocytes occupy it and isolate the neurons individually in their little cocoons. (In the periphery, e.g., in ganglia, this is exactly what the satellite cells do.)
Other Glial Elements
 Another group of glial cells of the CNS include the oligodendrocytes, which are
the equivalent of the Schwann cells of the peripheral nervous system. Like the Schwann cell, the oligodendrocyte has the job of making the myelin sheath around axons. Oligodendrocytes are inconspicuous cells, usually seen in Cresyl Violet/Luxol Fast Blue preparations as small, clear cells with a centrally located nucleus and a rim of unstained cytoplasm, rather like the yolk of an egg.
Another group of glial cells of the CNS include the oligodendrocytes, which are
the equivalent of the Schwann cells of the peripheral nervous system. Like the Schwann cell, the oligodendrocyte has the job of making the myelin sheath around axons. Oligodendrocytes are inconspicuous cells, usually seen in Cresyl Violet/Luxol Fast Blue preparations as small, clear cells with a centrally located nucleus and a rim of unstained cytoplasm, rather like the yolk of an egg.
The microglial cells of the CNS are phagocytic cells that protect neurons by engulfing potentially harmful materials. The ependymal cells of the spinal cord's central canal have already been mentioned.