Objectives for This Exercise
Laboratory Exercise on Hair
Slides for this Exercise: 24, 1205, 7, 11, 35, H9190, VM 45, VM46, FH132, VM43, MCO0039, MCW023
PRE-RECORDED LECTURES FOR THIS EXERCISE
Epidermis
Adnexa
Nervous Elements of Skin
One of the most difficult bits of mental gymnastics you will ever do is the visualization and reconstruction of 3-dimensional objects from 2-dimensional microscope slides. In working with the skin, you must do this for the skin itself, and for the accessory structures present in it. Structures such as glands and hair follicles look different at different levels, and two adjacent structures may well be sectioned at different levels in the same slide.
The integument of mammals consists of the skin proper, and also the various adnexa or accessory structures. These are things like hair, glands, nails, hooves, claws, and so forth. All of these are ultimately derived from the skin and are, in fact, part of it.
The epidermis is the epithelial portion, in which cellular proliferation and renewal takes place. Remember that epithelial sheets are avascular: the epidermis is no exception to this rule. No blood vessels run through it, though some run close by. Nor are nervous structures actually found in the epidermis proper. They are located in the next layer below it.
As with other epithelial sheets, the regenerative capacity of the epidermis is high. Rapid and continuous production of new epidermal cells in the deep portions is needed to keep up with the constant wearing away of cells on the surface. A typical human loses perhaps as much as 250 grams of skin a day, flaked off at the surface and renewed by mitosis in the deep epidermis. The accessory structures of the skin—the adnexa—such as hairs and glands, all epidermal derivatives, have similar capabilities of regeneration. But it's the epithelium that is turned over constantly under normal circumstances.
The dermis is the connective tissue layer underlying the epidermis. It's here you will find the blood vessels which serve the nutrient needs of the deep epidermis, as well as the nerve fibers that carry the signals from receptors located just under the epidermis. There are also lymphatic channels, etc. The dermis is a dense irregular collagenous CT, and, as with any such location, you'll find wandering CT cells (e.g., mast cells) in here. The dermis of well-nourished animals may have aggregations of adipocytes in it. The dermis is where sutures are actually anchored when closing a wound, since the epidermis is mechanically very weak. Coursing through it are numerous small blood vessels, with capillary loops forming close to the epidermal layer in various places. The cells of the avascular epidermis receive their nourishment and waste disposal services from the underlying dermis. Only the living layers near the boundary need be serviced, as the upper layers are dead and sloughing off.
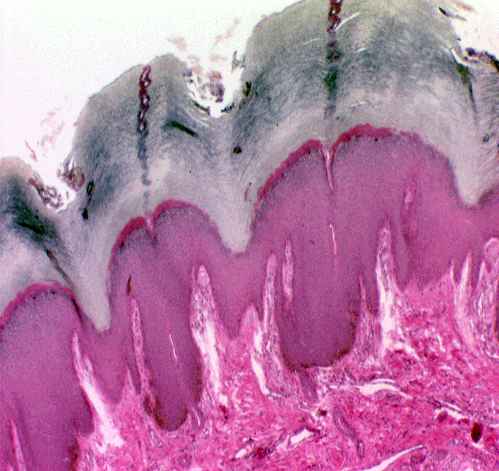
 Technically not part of the skin, but closely associated with it is the hypodermis or "subcutis."
This is a layer of connective tissue that lies deep to the dermis, as its name
implies. The hypodermis is a somewhat looser but still irregular CT. In animals
in good condition it will have large depots of fat in it. Blood vessels course
through here on their way to and from the dermis. When you skin an animal this
is the "loose fascia" that's under the skin—the separation of the
layers usually is at the dermo-hypodermal boundary, which is pretty indistinct
in a microscope.
Technically not part of the skin, but closely associated with it is the hypodermis or "subcutis."
This is a layer of connective tissue that lies deep to the dermis, as its name
implies. The hypodermis is a somewhat looser but still irregular CT. In animals
in good condition it will have large depots of fat in it. Blood vessels course
through here on their way to and from the dermis. When you skin an animal this
is the "loose fascia" that's under the skin—the separation of the
layers usually is at the dermo-hypodermal boundary, which is pretty indistinct
in a microscope.
It's in or near the hypodermis that most of the accessory structures (see below) are seen, although as epithelial structures and epidermal derivatives, they're separated from the dermis and hypodermis proper by a basement membrane.
The image at right is from slide 1205. The hypodermis is usually much more heavily infiltrated with fat than the dermis, and there can be very large fat depots in it in some species, especially when they're well-fed. If you've ever seen a skinned pig carcass, you're looking at about 4" of hypodermis after the dermis and epidermis have been removed. A "hypodermic injection" is one that goes below the dermis and into the hypodermis to deliver the drug.
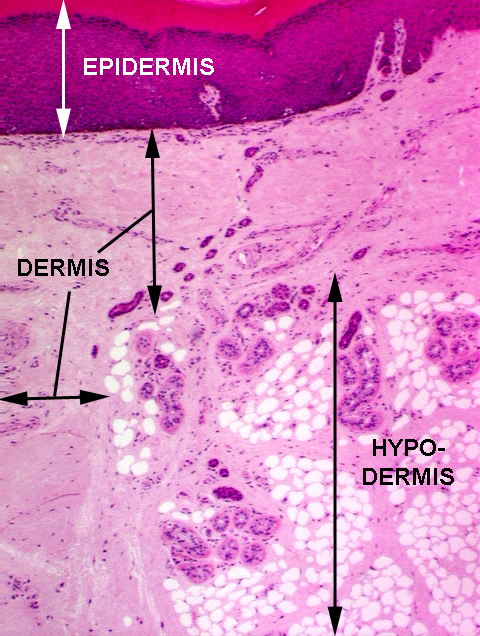
 alongside a drawing of a similar section stained with the Mallory connective tissue stain. This will clarify the relationship of the fibrous CT and the fat depots. Notice the numerous capillaries present in these images. The actual boundary between the dermis and the hypodermis isn't usually all that clearly demarcated in histological preparations, especially of thin skin (see below) but in this section from a footpad, the fat depots are large, the CT pretty dense, and you can label anything with a depot the size of this one as "hypodermis" very easily.
alongside a drawing of a similar section stained with the Mallory connective tissue stain. This will clarify the relationship of the fibrous CT and the fat depots. Notice the numerous capillaries present in these images. The actual boundary between the dermis and the hypodermis isn't usually all that clearly demarcated in histological preparations, especially of thin skin (see below) but in this section from a footpad, the fat depots are large, the CT pretty dense, and you can label anything with a depot the size of this one as "hypodermis" very easily. The vast majority of cells in the skin are keratinocytes. These cells are produced by constant mitosis in the deeper regions, move rapidly up to the surface, die, are worn away, only to be replaced by the next wave of newly-produced cells. The life of a given keratinocyte is about 7 to 10 days from production to sloughing. These are the components of the "keratinzing system." Keratinocytes make up about 95% of the total number of cells in the epidermis.
For now we'll be concerned solely with keratinocytes and their fate, and look at other cell types (not part of the keratinizing system) later. The epidermis has several distinct layers, all in well-defined stages of keratinocyte life history. The keratinocytes begin as viable, dividing cells, and end as nothing more than flattened, battered, dead sacs of keratin. You can identify the different regions of the epidermis on slide 1205.
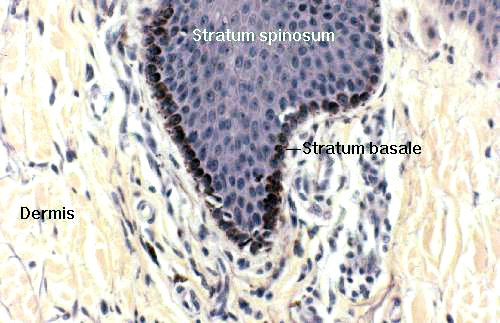
 The deepest layer of the epidermis is the stratum basale. This is a
single layer of keratinocytes, those right up against the dermo-epidermal
boundary. It's here that almost all of the proliferative activity occurs. This
layer is only one cell deep, by definition.
Any agent, chemical or physical, which interferes with cell division will result in sloughing of the skin because the replacement cycle is affected. One of the very early symptoms of radiation overexposure is skin loss. Cytotoxic drugs used in cancer therapy have a similar effect, because their target is rapidly proliferating cells.
The deepest layer of the epidermis is the stratum basale. This is a
single layer of keratinocytes, those right up against the dermo-epidermal
boundary. It's here that almost all of the proliferative activity occurs. This
layer is only one cell deep, by definition.
Any agent, chemical or physical, which interferes with cell division will result in sloughing of the skin because the replacement cycle is affected. One of the very early symptoms of radiation overexposure is skin loss. Cytotoxic drugs used in cancer therapy have a similar effect, because their target is rapidly proliferating cells.
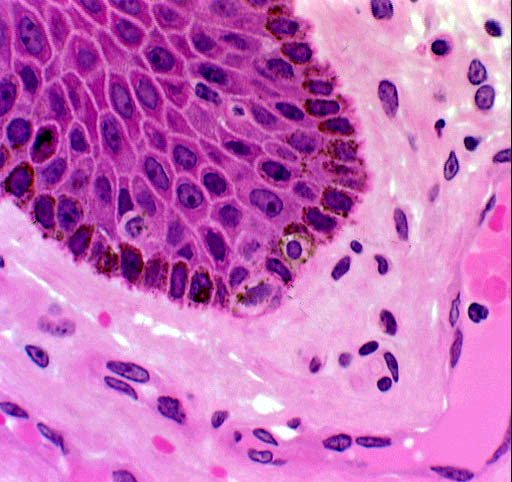
It's obvious from the image shown here that there's more than one cell type in the stratum basale. We'll look at the details of this later, but the unstained cells in this picture are melanocytes, which make up most of the rest of that 5% that isn't keratinocytes. Notice also that there's a blood vessel (in this case a venule) just below the dermo-epidermal boundary; and numerous cells of the dermal CT.
Stratum Spinosum of the Epidermis
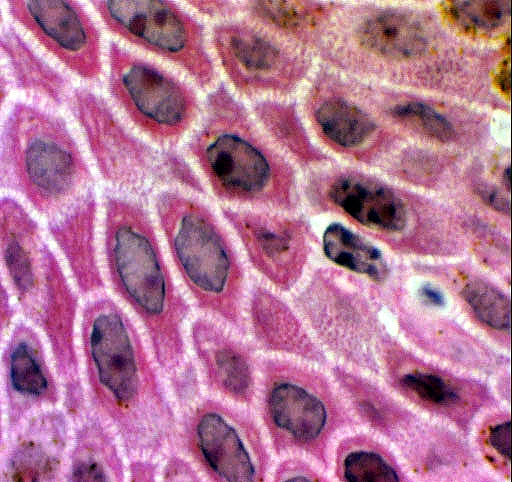 Moving one level up from the stratum basale, you'll find the stratum spinosum,
sometimes called the "prickle cell" layer. In this layer the cells
are still alive, at least in the deeper regions of it. they can even divide, to a limited extent, in cases of high demand for new keratinocytes. As they creep upward, however, these cells lose their capability for division and they make the transition from living to barely-living, and eventually to dead.
Moving one level up from the stratum basale, you'll find the stratum spinosum,
sometimes called the "prickle cell" layer. In this layer the cells
are still alive, at least in the deeper regions of it. they can even divide, to a limited extent, in cases of high demand for new keratinocytes. As they creep upward, however, these cells lose their capability for division and they make the transition from living to barely-living, and eventually to dead.
The stratum spinosum has a peculiar appearance in LM preparations: there are what appear to be spines or "hairs" protruding out from them, and connecting them to adjacent cells. With high magnification you'll be able to make out these structures, historically called "intercellular bridges" which are characteristic of this layer. In fact these "intercellular bridges" are an artifact of preparation, causes by shrinkage. Nevertheless, they're so consistently present that they can reliably be used as an identifying characteristic of this layer.
Keep in mind, however, that there is no continuity of cytoplasm between adjacent cells. The "bridges" are sites where the adjacent cells of the stratum spinosum are held firmly together by desmosomes. The desmosomes can't be seen with a light microscope, but post-mortem and post-fixation shrinkage causes stretching of cellular material at the points of adhesion, creating the illusion of "bridges." The result is the ladder-like appearance in the intercellular space surrounding adjacent keratinocytes. The true nature of these wasn't appreciated until about half a century ago, when the electron microscope made it possible to examine their architecture at high resolution. The stratum spinosum may be quite extensive in thick skin, but in thin skin it may be only one or two cells' breadth.
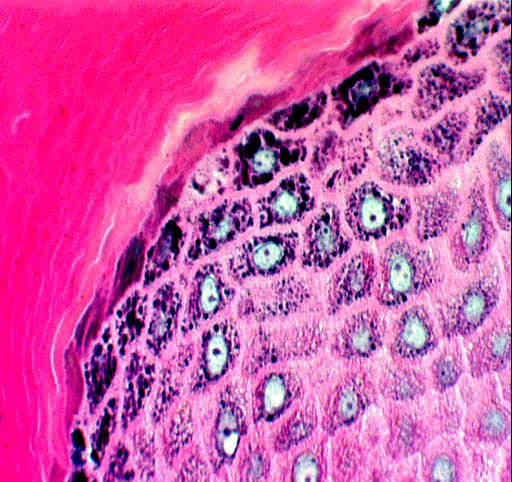 Moving again upwards, the next layer is the stratum granulosum. This
layer takes its name from the granular pre-keratin material usually seen in its
keratinocytes. This material takes the form of aggregated keratohyaline granules. Cells of the stratum granulosum are dead and can no longer divide.
Moving again upwards, the next layer is the stratum granulosum. This
layer takes its name from the granular pre-keratin material usually seen in its
keratinocytes. This material takes the form of aggregated keratohyaline granules. Cells of the stratum granulosum are dead and can no longer divide.
The stratum granulosum isn't uniformly seen in sections. It's usually found in thick skin without trouble, but in thin skin, it may be attenuated to one or two cells or even missing altogether. You may sometimes read that "wet keratinzed squamous epithelium doesn't form a stratum granulosum; dry keratinzed epithelium does." THIS IS NOT TRUE. It's possible to find a stratum granulosum in, for example, the keratinized stratified squamous epithelium of the oral cavity and/or esophagus in many species. Keratin is a fibrous protein, the stuff of fingernails, claws, and hair (all epithelial derivatives) but exactly what the granules are for or what they represent is something of a mystery. They're thought to be a keratin precursor, but their presence in keratinocytes isn't required for keratinization to occur, and their presence isn't a guarantee that keratinization is inevitable.
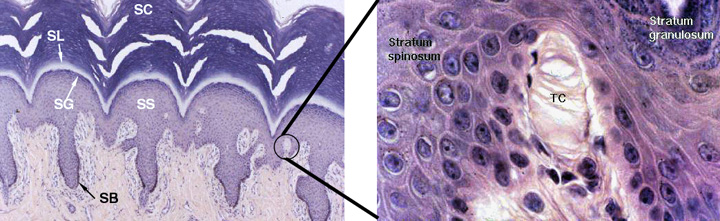
This is another "optional" epidermal layer: you won't find it in thin skin, but it's a feature of thick skin, especially in places of very heavy wear and consequently heavy keratinization, such as foot pads. It isn't formed in regions of skin which have hair follicles. You'll see it as a clear band above the stratum granulosum. "Lucidum" is Latin for "clear," hence the name of this layer based on its appearance in microscopic preparations.
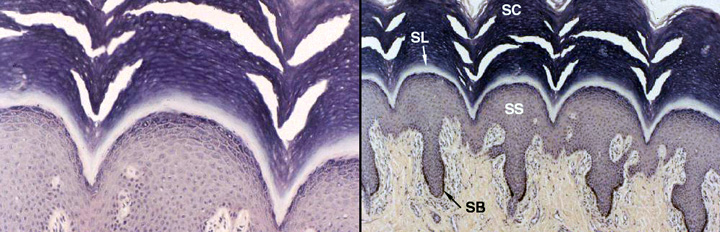
The cells in this layer are deader than the Pharaohs, lifeless bags of plasma membrane filled with eleidin, another fibrous protein. It may be an intermediate form of keratin, a transitional molecule between the keratohyaline granules' substance and the fully-formed keratin fibers in the next layer, the stratum corneum. The image above is from slide 24, a very heavily keratinized specimen. The stratum lucidum is obvious. The stratum spinosum (SS) and stratum basale (SB) are labeled (note the melanin in the latter!) and the stratum granulosum (not labeled) is visible in the left-hand image. The stratum corneum (SC) is visible at the top of the fields.
At the surface, the outermost epidermal layer is found. This is the stratum corneum
or "horny layer." It's the actual
interface between the outer surface of the animal and its environment, the layer from whence cells are cast off in the normal process of wear and tear. The very outermost portion of the stratum corneum is constantly in the process
of sloughing off; from time to time you may see a reference to the
"stratum disjunctum," and that's what is meant by the term. Socially
speaking, it's incipient dandruff.
In this image from slide 24 you can see the corkscrew-shaped channels of sweat glands (see below) working their way up from the deep layers of the skin, through the very heavy stratum corneum, and eventually opening at the surface.
Thin Skin vs Thick Skin
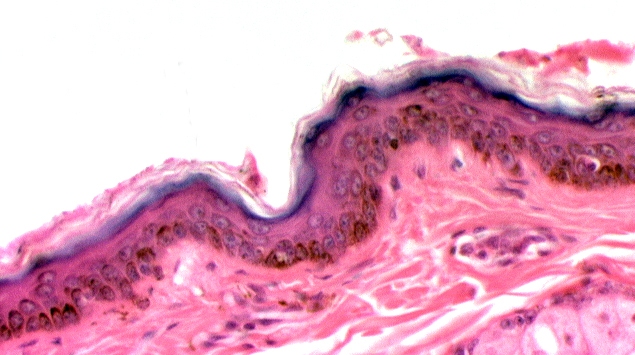 We've been mainly discussing features of thick skin. Thin skin differs from thick in a few ways, but mainly in the lack of a stratum
lucidum. The stratum granulosum is often absent or discontinuous. Examine slide
7, a section of thin skin from a pig.
We've been mainly discussing features of thick skin. Thin skin differs from thick in a few ways, but mainly in the lack of a stratum
lucidum. The stratum granulosum is often absent or discontinuous. Examine slide
7, a section of thin skin from a pig.
The stratum corneum is thin, and a stratum disjunctum is present. There is no stratum lucidum below it, but in places you may see a scanty (1-2 cell layer thick) stratum granulosum, and in places it's absent. The stratum spinosum is present, as it always is, but again is only a couple of cells thick, and deep to that is the stratum basale. In this deepest part of the epidermis you will find that mitotic activity is quite high, and you should be able to identify some cells in mitosis.
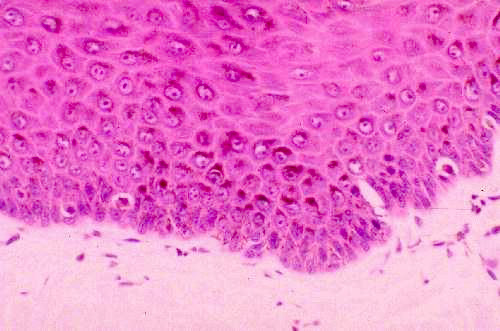 While keratinocytes are by far the most numerous cells in the epidermis, they
aren't the only type. The pigmentation in skin comes partly from melanocytes,
another very important cell. Slides 24 and 1205 show melanocytes quite clearly.
In the regions where the brown melanin pigment is concentrated, look in among
the cells of the stratum basale for cells which are unstained.
While keratinocytes are by far the most numerous cells in the epidermis, they
aren't the only type. The pigmentation in skin comes partly from melanocytes,
another very important cell. Slides 24 and 1205 show melanocytes quite clearly.
In the regions where the brown melanin pigment is concentrated, look in among
the cells of the stratum basale for cells which are unstained.
Melanin, the most common of the biological pigments, is produced by special cells derived from the neural crest and located in specific places. One such place is the deep layers of the skin.
In this H&E preparation melanin appears as brownish material inside the cells of the skin's deep layers. The cells which produce it (melanocytes) are residents of these deep regions and pass the pigment on to other cells that carry it to the surface. Melanin gives skin part of its coloration, and the inability to synthesize it is the condition we call albinism; an albino animal has white fur or feathers and no pigmentation in its eyes, either.
 The reverse condition, sometimes seen as a color phase variation in rodents and carnivores, is melanism. In this condition more than the usual amount is made. Such animals are said to be melanistic. The famous black squirrels found in the streets and parks of Washington DC are a nice example. They're melanistic strains of the common grey squirrel (Sciurus carolinensis). these attractive animals were introduced to the National Zoo several times before World War One, from Rondeau Provincial Park in Ontario. Melanistic squirrels are far more common in northern climates, where the dark color is thought to help them absorb radiant heat in the very cold winters. Albino squirrels aren't nearly so common, but they do exist: the town of Olney, Illinois has a large population of them, and has adopted the white squirrel as the town's official symbol! Despite their appearance they're the same species, they freely interbreed with "normal" greys. In a mixed area S. carolinensis can display a wide range of color variations from white, to normal grey, to a beautiful glossy, deep blue-black.
The reverse condition, sometimes seen as a color phase variation in rodents and carnivores, is melanism. In this condition more than the usual amount is made. Such animals are said to be melanistic. The famous black squirrels found in the streets and parks of Washington DC are a nice example. They're melanistic strains of the common grey squirrel (Sciurus carolinensis). these attractive animals were introduced to the National Zoo several times before World War One, from Rondeau Provincial Park in Ontario. Melanistic squirrels are far more common in northern climates, where the dark color is thought to help them absorb radiant heat in the very cold winters. Albino squirrels aren't nearly so common, but they do exist: the town of Olney, Illinois has a large population of them, and has adopted the white squirrel as the town's official symbol! Despite their appearance they're the same species, they freely interbreed with "normal" greys. In a mixed area S. carolinensis can display a wide range of color variations from white, to normal grey, to a beautiful glossy, deep blue-black.
 Melanocytes make the melanin, but they don't retain it for long. Instead, they
pass it to the neighboring keratinocytes which then carry it up through the
layers and out of the system. Although you can't see them in the LM,
melanocytes have long and extensive processes that reach out and contact many
keratinocytes, to facilitate pigment transfer. In places where melanocytes are
concentrated and/or very active, localized deep pigmentation occurs.
Melanocytes make the melanin, but they don't retain it for long. Instead, they
pass it to the neighboring keratinocytes which then carry it up through the
layers and out of the system. Although you can't see them in the LM,
melanocytes have long and extensive processes that reach out and contact many
keratinocytes, to facilitate pigment transfer. In places where melanocytes are
concentrated and/or very active, localized deep pigmentation occurs.
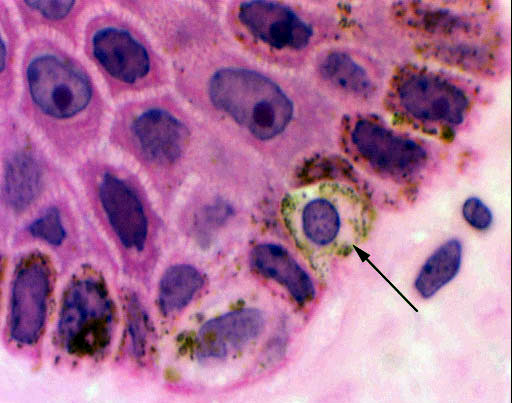 The melanocytes, unlike the keratinocytes, are a very stable population of
cells. They normally do not divide, and their life span is measured in years to
decades, rather than the few days of the keratinocytes. If, however, they
do "decide" to divide, the consequences are usually serious. A melanoma
is a type of very aggressive and metastatic cancer which arises from the
uncontrolled multiplication and migration of these cells.
The melanocytes, unlike the keratinocytes, are a very stable population of
cells. They normally do not divide, and their life span is measured in years to
decades, rather than the few days of the keratinocytes. If, however, they
do "decide" to divide, the consequences are usually serious. A melanoma
is a type of very aggressive and metastatic cancer which arises from the
uncontrolled multiplication and migration of these cells.
Two more cell types are found in the epidermis, and we'll note them here for the record, though you probably won't be able to recognize them in your slides. Neither of them constitutes more than a small fraction of the total number of cells in the epidermis. They're best seen with special stains, but if you happen to come across an "unstained" cell that's clearly in the stratum spinosum, it will be one of these two.
The Merkel cell is found in small numbers near the stratum basale and appears to be most numerous in places with special sensitivity, such as the fingertips. They form communications with local neurons that suggest they have a neurosensory function.
The Langerhans cell is found in the upper stratum spinosum and it's part of the immune system. These cells have surface markers for recognition by T-lymphocytes, and it's believed they take up antigens and present them to the T-lymphocytes for response. They're an important participant in allergic responses.
The epidermis is not a flat sheet. Its border with the underlying connective tissue of the dermis is an undulating and irregular line. Downward projections of epidermis (epidermal pegs) are matched to corresponding upthrusts of the dermis (dermal ridges) forming an interlocking boundary. This anchors the epidermis in place. The degree of interdigitation is related to the level of wear; areas with high levels of wear have lots of interdigitation, while areas of low wear have little (though there is always some). These pegs and ridges actually represent "hills and valleys" cut in cross section or oblique section. If you want to see what they look like in three dimensions, get a magnifying glass and look at your fingertips. The interdigitation of dermal ridges and epidermal pegs is what makes the unique pattern of your fingerprints.
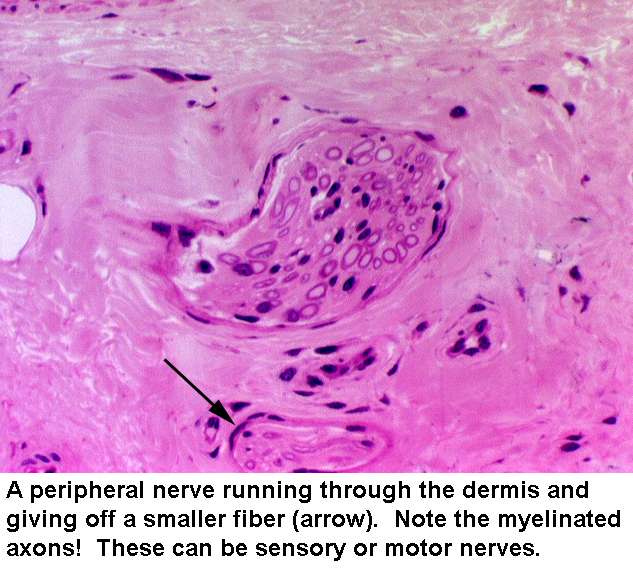 Many nerve fibers run through the dermis, carrying messages from sensory
receptors at the periphery of the body. At left is an example of peripheral nerve in the dermis of the skin. Get into the habit of looking for these and for blood vessels in the dermis of skin preparations. Keep in mind that you can see them cut at any angle from perfect cross sections to perfect longitudinal sections and anything in between. Myelin sheaths should be visible, and in longitudinal sections, you may see nodes of Ranvier.
Many nerve fibers run through the dermis, carrying messages from sensory
receptors at the periphery of the body. At left is an example of peripheral nerve in the dermis of the skin. Get into the habit of looking for these and for blood vessels in the dermis of skin preparations. Keep in mind that you can see them cut at any angle from perfect cross sections to perfect longitudinal sections and anything in between. Myelin sheaths should be visible, and in longitudinal sections, you may see nodes of Ranvier.
Inside the dermal papillae, close up under the epidermis, you'll also find some examples of the skin's sensory transducer structures. There are dozens of varieties (receptors for heat, cold, pain, pressure, etc.), but they tend to run in a pattern anatomically, and we'll only look at a couple that typify them all. As a class, they're transducers. Their role is to take whatever stimulus they're adapted to detect and convert that into a signal the nervous system can perceive. The signal then is carried via a sensory nerve fiber to the CNS (perhaps the one shown above) and decisions are made there as to what to do about the information.
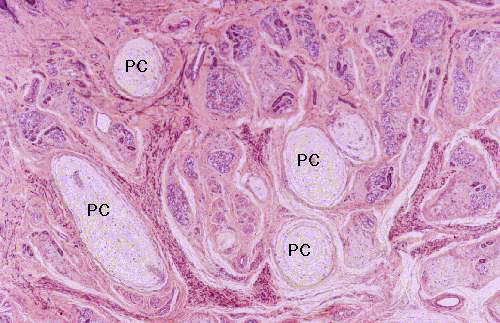
On slide 1205, you'll be able to locate some tactile corpuscles. These are transducer structures; they're closely associated with neural fibers and serve to change the mechanical signal of pressure into a neuronal impulse.
The tactile corpuscle (TC) in this field is located in the dermis, though it's as close to the epidermis as one can get! It's right up against the boundary of the two layers. Look inside the dermal ridges on slide 24, where the interdigitation is great, and you'll see them. This one's a very nice example of the whole class of peripheral mechanical transducers, an elongated, club-shaped pile of CT, butted up against the dermo-epidermal boundary. There's a nerve fiber in it (though it isn't visible in this preparation), an afferent fiber from a pseudo-unipolar neuron in a sensory ganglion. Pressure on the transducer exerted along its long axis deforms the corpuscle.
The nerve fiber fires because mechanical deformation affects its membrane charge, and a signal is sent. Sensory corpuscles for various sensations (heat, cold, touch, etc.) are known to exist, but anatomically they are pretty much indistinguishable. They are (as you would expect) most numerous in those parts of the skin where touch is most sensitive, as for example the fingertips of primates.
Tactile corpuscles are arranged so that their long axis is perpendicular to the skin's surface. When pressure is placed on them, the stack is compressed, and the nerve fiber physically distorted. This changes the surface membrane's charge characteristics, and the fiber sends a signal to it's neuron, which then duly relays the information to the CNS for further action. Virtually all of the mechanical transducers of the skin use some variant of this mechanism.
Lamellated (Pacinian) Corpuscles
 In the deeper parts of this slide, i.e., the hypodermis, you will find a lamellated
corpuscle. The ones shown here are from slide 1205, but you may find them on slide 24 as well. These are also called "Pacinian corpuscles" after their discoverer, Filipo Pacini. You can hardly miss this structure: it's very large, and looks much like a sliced onion in sections. If you are fortunate you will be
able to see the nerve fiber which penetrates the "onion" to the
innermost layer. These corpuscles can be big enough to see with the naked eye: some of them are the size of pencil point.
In the deeper parts of this slide, i.e., the hypodermis, you will find a lamellated
corpuscle. The ones shown here are from slide 1205, but you may find them on slide 24 as well. These are also called "Pacinian corpuscles" after their discoverer, Filipo Pacini. You can hardly miss this structure: it's very large, and looks much like a sliced onion in sections. If you are fortunate you will be
able to see the nerve fiber which penetrates the "onion" to the
innermost layer. These corpuscles can be big enough to see with the naked eye: some of them are the size of pencil point.
The lamellated corpuscle works the same way the tactile corpuscle does but on a much grosser and coarser scale. The
layers of CT are physically 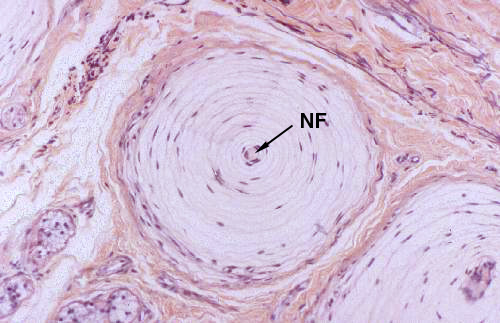 compressed and the shape is distorted. This
mechanical pressure causes the nerve fiber inside to depolarize and fire,
sending a signal to the central nervous system. The lamellated corpuscle is a
much coarser transducer than the tactile corpuscle is. Lamellated corpuscles
are gross pressure receptors, and/or vibration receptors. (They're found in
visceral organs, too. That interesting feeling you experience when you are in a
very fast elevator—you know, when you go down swiftly and your stomach is two
floors above you?—comes from input sent to the brain by lamellated
corpuscles.)
compressed and the shape is distorted. This
mechanical pressure causes the nerve fiber inside to depolarize and fire,
sending a signal to the central nervous system. The lamellated corpuscle is a
much coarser transducer than the tactile corpuscle is. Lamellated corpuscles
are gross pressure receptors, and/or vibration receptors. (They're found in
visceral organs, too. That interesting feeling you experience when you are in a
very fast elevator—you know, when you go down swiftly and your stomach is two
floors above you?—comes from input sent to the brain by lamellated
corpuscles.)
Sensory receptors exist for cold and heat, and other sensations as well. They look much like the tactile corpuscles. The receptor for pain (euphemistically called a "nociceptor" from Latin, noceo, to injure or hurt) is often a naked nerve fiber ending. These will require special stains to be seen.
Accessory structures present on this slide include hairs and two types of glands: sweat glands and sebaceous glands. Although most of these structures are "anchored" in the dermis or hypodermis, they are in fact all of them in and of the epidermis. All of the skin's adnexa are epidermal in origin, and careful examination (especially with slides made with CT stains) will reveal that hair follicles and sweat glands, sebaceous glands, etc. always are separated from the underlying dermal tissue by a basal lamina. They truly are in the epidermis.
The sweat glands are the most conspicuous accessory structures in slides 7, 35, and 1205; they are seen as hollow, more or less circular profiles, with walls composed of a very low cuboidal epithelium, and lumens that appear to be empty. The sweat glands' peculiar appearance in sections results from their overall structure as simple coiled tubular glands. In passing the plane of the cut through the coil of the sweat gland, it's cut in several places. If you were to slice through a coiled garden hose, you would get much the same result—you would be able to look into the cut portions of the hose in several places.

The drawing at the left shows an idealized representation of the sweat gland as seen in sections: an actual specimen is shown next to it, with the center image showing mainly the duct region; the right image shows secretory cells and the deepest region of the duct. Note the fat cells in the nearby CT: 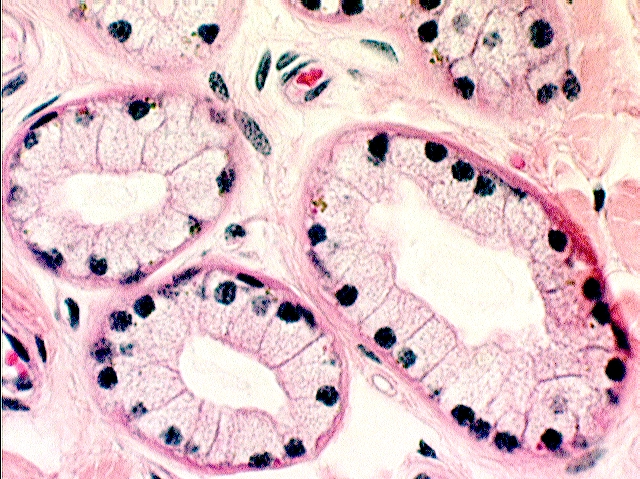 the deep region of the gland is surrounded by hypodermis, and the duct worms its way upwards, towards the surface, eventually leading the secretion to the outside. This gland is of the eccrine type (see below).
the deep region of the gland is surrounded by hypodermis, and the duct worms its way upwards, towards the surface, eventually leading the secretion to the outside. This gland is of the eccrine type (see below).
If you examine the secretory portion at high magnification you'll find thin boundary cells resembling smooth muscle demarcating the limits of the secretory portions. The secretory parts themselves are composed of simple cuboidal epithelium, and the lumen of one of these glands often appears patent.
Two types of sweat glands exist: small eccrine sweat glands that produce a watery secretion, and larger apocrine sweat glands whose secretions are more oily and foamy. The ones shown above are eccrine type, mainly from slides 7, 11, 12, and 35. Eccrine sweat glands can be found on all the slides looked at so far, and also on slide 12, the scrotal skin of a dog. The secretory regions lie deep, and the ducts make their way to the surface. On slide 24, if you look at the stratum corneum, you should be able to make out the opening of the ducts as they corkscrew up through to the surface. Dogs do sweat (no matter what you have heard) and on extremely hot days may actually leave damp pawprints.
These small glands are widely distributed: they're mainly a mechanism for cooling. The perspiration they produce evaporates and takes heat with it. Since perspiration is under nervous control (the CNS is aware of the ambient temperature, of course: beta-adrenergic nerve fibers innervate the eccrine glands), drugs which have the effect of blocking "beta receptors" have as one of their side effects the diminution of sweating. This can actually be dangerous if the individual is subjected to extremely hot weather and can't sweat off some of the heat load.
Apocrine Glands
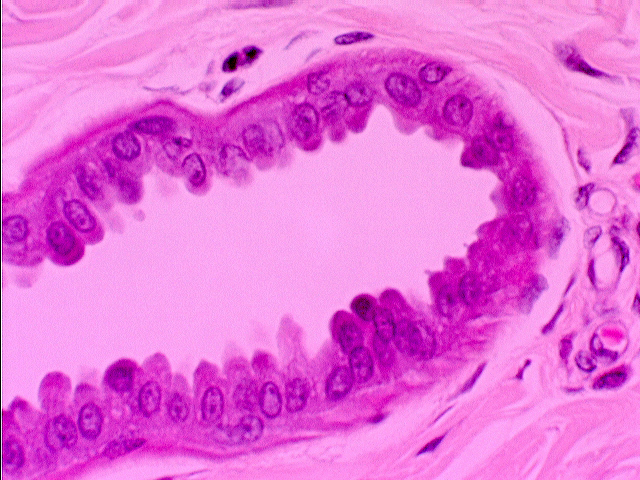 The apocrine sweat glands are probably best seen on slide 35. These are
more limited in distribution than the eccrine glands. They're typically found in the groin, the axilla, and
on the scrotum. Horses have lots of them, and they're more widely distributed
in equines than in other groups. If you ride a horse till he's "lathered up,"
the "lather" is the sweat of the eccrine glands plus the oily output
of these very large apocrine glands, which gives it the foamy appearance it
has.
The apocrine sweat glands are probably best seen on slide 35. These are
more limited in distribution than the eccrine glands. They're typically found in the groin, the axilla, and
on the scrotum. Horses have lots of them, and they're more widely distributed
in equines than in other groups. If you ride a horse till he's "lathered up,"
the "lather" is the sweat of the eccrine glands plus the oily output
of these very large apocrine glands, which gives it the foamy appearance it
has.
Look deep in the dermis for profiles similar to those of the eccrine glands, but much larger. Note that in the lumina of these glands, you can see a sort of "pinching off" effect, where the apical cytoplasm seems to be coming off into the lumen. It is. The secretion of these glands consists of parts of the cytoplasm, pinched off, cast away, lysed, and necrotized.
Another common feature of the skin, yet another gland type, is the sebaceous gland. We'll look at these in more detail in Exercise 15, dealing with hairs and hair follicles.