Objectives for This Exercise
Laboratory Exercise on Skin
Slides for this Exercise: 24, 1205, 7, 11, 35, H9190, VM 45, VM46, FH132, VM43, MCO0039, MCW023
True hair is found only in the Class Mammalia, and there is really no such thing as an absolutely hairless mammal. Even whales (at least some of them) have rudimentary hairs here and there. Some other animals have hair-like structures, but if you have real hair, you're a mammal.
Hair serves many functions. The most obvious is to serve as insulation, but it's also used to provide camouflage (for example, the spot pattern on a fawn deer mimics the play of light and shade on a forest floor), for sex recognition (male mammals often have ruffs, manes, or beards as secondary sex characteristics) and even for social purposes, such as aggressive display (as when a dog's "hackles rise" or a cat elevates her fur on the approach of a dog).
Compare the diagram above to the low magnification view of an active hair follicle cut almost perfectly along the longitudinal axis. In addition to the hair proper, various structures that are part of the follicle are visible, including a sebaceous gland, a sweat gland, and the arrector pili muscle (APM). The hair bulb is that portion of the follicle which actively produces the hair; the root sheath(s) surround the hair as it grows out. All hairs, of whatever species, and regardless of location on the body, grow out of follicles much like this one. We'll look at each of these components of the follicle in detail below.
Start with slide 11, the nose of a dog. This slide has a fair number of large hairs on it, and you will be able to identify the principal parts by referring to these two images. Look for a place where a sebaceous gland opens into the follicle; you should also be able to see numerous profiles of sweat glands also visible near the base of the hair. Some sweat glands, especially in thin integument, also open into the follicles, some don't.
The Hair Follicle Is Epithelial In Nature
The hair grows from the hair follicle, not out of it. One point that needs to be emphasized is that the hair follicle and the hair it produces are part of the epidermal region of the skin. The follicle is separated from the underlying dermis and hypodermis by a basement membrane, and the hair itself grows from the germinal epithelium at the deepest part of an active follicle. Hairs don't grow "through" the skin. They grow from skin and in a very real sense they're part of the keratinizing system of the epidermal layer.
 The arrector pili muscle associated with each follicle is a band smooth
muscle whose origin is in the fibrils of the dermis. It inserts onto the
CT investment of the follicle. Unlike
the hair, which is ectodermal in origin, and part of the epidermis, the arrector pili is a true dermal
structure. Muscle is derived from embryonic mesoderm, and the arrector pili is no exception. The collagen fibers of the dermis are also mesodermal.
The arrector pili muscle associated with each follicle is a band smooth
muscle whose origin is in the fibrils of the dermis. It inserts onto the
CT investment of the follicle. Unlike
the hair, which is ectodermal in origin, and part of the epidermis, the arrector pili is a true dermal
structure. Muscle is derived from embryonic mesoderm, and the arrector pili is no exception. The collagen fibers of the dermis are also mesodermal.When the arrector pili contracts, it elevates the hair. This phenomenon of piloerection can occur in response to a number of stimuli. Piloerection is a common response to cold; lofting the hair increases the air spaces and hence the insulating value of the hair coat. It also occurs in response to anger or fear, because of the adrenaline surge that these emotions involve. If you've ever seen two dogs who've taken a dislike to each other getting ready to settle their differences, you will have witnessed a nice demonstration of this. It makes plain why we speak of a dog "bristling" at another, and why we say "his hackles rise" and he "has his back up." Cats do the same thing when confronted by dogs, as a desperate (and often successful) attempt to make themselves look bigger and more formidable. Humans have only a vestigial hair coat (Desmond Morris titled his book on human behavior The Naked Ape for good reason; it's an apt description), but we do the same thing. The contraction of our arrector pili muscles causes "goosebumps."
Compound Follicles

At left is a cross section through a compound follicle. This is a follicle from which more than one hair emerges. Typically there's a large main guard hair and several smaller wool hairs with small or absent medullary regions (see below). All the hairs usually emerge from the same opening on the surface. The guard hair provides a waterproof top layer and the curly undercoat, made of wool hairs, an insulation barrier against the skin. Northern-breed dogs and other animals that live in wet, cold, climates have this type of hair coat.
Sebaceous Glands
Notice in this slide also the presence of sebaceous glands. This adnexal structure's function is to provide lubrication and waterproofing to the hairs. The sebaceous gland is sort of an extreme version of the apocrine sweat gland. In apocrine glands, cytoplasm is pinched off and cast into the lumen, where it disintegrates and becomes the secretion. Sebaceous glands are the only example in mammals of a holocrine gland, one in which the entire cell body of the glandular cells is sacrificed to become the secretory material.
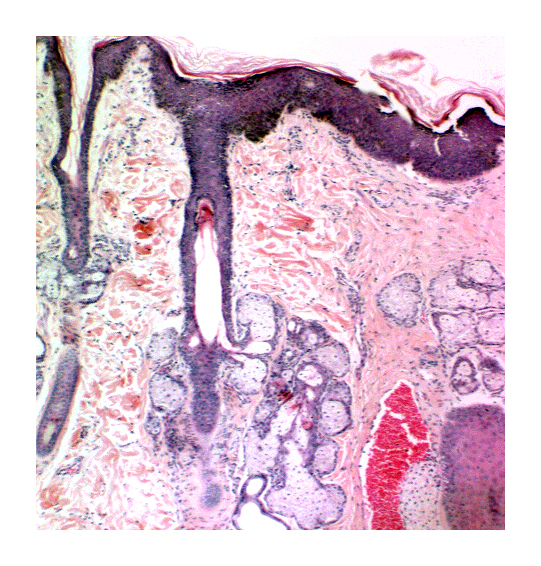 At right is a follicle cut longitudinally, with a large array of sebaceous gland units visible in the section. The sebaceous gland is simple branched acinar in structure: all the pear-shaped secretory units discharge into a single short duct, that leads into the hair follicle about halfway towards the surface, just about at the level where the inner root sheath disappears. The sebaceous gland engages in holocrine secretion. The lysed cells of its substance, cast off into the lumen and allowed to ooze out onto the surface of the skin.
At right is a follicle cut longitudinally, with a large array of sebaceous gland units visible in the section. The sebaceous gland is simple branched acinar in structure: all the pear-shaped secretory units discharge into a single short duct, that leads into the hair follicle about halfway towards the surface, just about at the level where the inner root sheath disappears. The sebaceous gland engages in holocrine secretion. The lysed cells of its substance, cast off into the lumen and allowed to ooze out onto the surface of the skin.
In this section, you can also see the continuity of the outer root sheath of the follicle (see below) with the surface epidermal layer. The small follicle on the left side of the field is in catagen and its hair has fallen out.
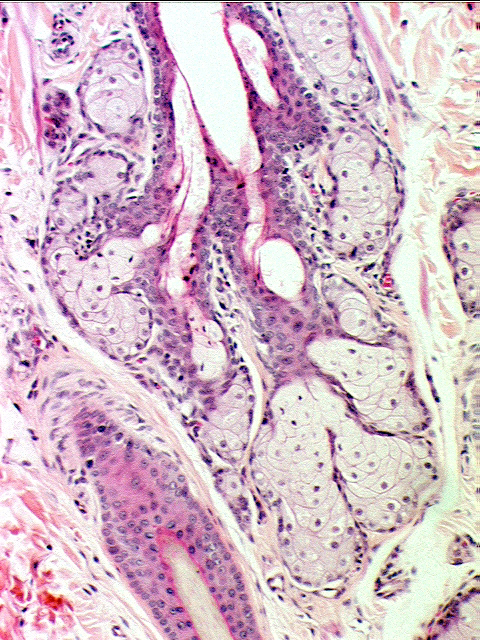 At higher magnification, you can see sebaceous gland units in some more detail. Note the "foamy" appearance of the cells in the sebaceous gland, and note also that the cells at the periphery of the gland are clearly healthy and alive, with normal nuclei. But if you look closer to the center of the gland, the nuclei become pyknotic, and then fragmented. The cells here are dying, and undergoing post-mortem lysis and disintegration to become the secretion.
At higher magnification, you can see sebaceous gland units in some more detail. Note the "foamy" appearance of the cells in the sebaceous gland, and note also that the cells at the periphery of the gland are clearly healthy and alive, with normal nuclei. But if you look closer to the center of the gland, the nuclei become pyknotic, and then fragmented. The cells here are dying, and undergoing post-mortem lysis and disintegration to become the secretion.
The foamy look comes from the very high lipid content of the cells. As the cells disintegrate, these lipids undergo breakdown, become rancid and separate into hydrocarbon chains of random lengths. This stuff is sebum. Discharging this goop into hair follicles gives hair its "sheen," acts as a lubricant for the skin and hair, and confers waterproofing and resistance to desiccation. Anything which affects the secretion of sebaceous glands—disease, drugs, etc.—will be reflected in the look and condition of the hair coat.
The particular combination of lipid breakdown products is unique to specific individuals. The scent signature that results is therefore also specific to the individual, and that's how your dog (whose nose is much better than yours) can tell you from everyone else he's ever met. Just as we visually-oriented animals never forget faces, animals that are olfactorily oriented never forget scent signatures.
Outer Root Sheath
Beginning at the surface of the skin to one side of the hair, follow the invagination of the epithelium down to the root (see image below). This invagination forms the outer epithelial root sheath, which you will see is continuous with the surface epidermis. The upper part of the sheath is usually keratinized, although the lower part generally isn't.
At the bottom of the hair follicle the epithelium of the outer sheath fuses with that of the inner sheath, and if the follicle is active, with the germinal matrix. The inner sheath is formed from the periphery of the germinal matrix; the hair is produced by the central region of the germinal matrix. Unlike the hair the inner sheath doesn't keratinize, and it never protrudes from the follicle opening. The inner sheath usually ends somewhere around the point of input of the sebaceous glands. The inner sheath has a couple of layers; on slide 35 you should be able to make out Huxley's layer (see below). It will nearly impossible to make out the cuticle, as it's very thin and applied closely to the hair.
The Hair Proper
 Let's look at the hair itself. In examining these slides, remember you're looking at a long, three-dimensional object, which could be cut at any angle from 0 to 90 degrees with respect to its long axis.
Let's look at the hair itself. In examining these slides, remember you're looking at a long, three-dimensional object, which could be cut at any angle from 0 to 90 degrees with respect to its long axis.
In cross sections (mainly slide 35) not all the hairs are cut on the same level, and some of the sheaths which end before the surface is reached will not be seen if you happen to have a section that is too high up. Use the presence or absence (and the size) of sebaceous glands as a rough indicator of where you are with respect to the "deep" or "shallow" parts of the follicle.
The one at left is from slide 11. This longitudinal section shows the outer root sheath and the cortex and medulla of the hair shaft very well; and the inner root sheath can be made out too. The inner root sheath doesn't go all the way up the side of the hair follicle: it ends about halfway to the surface. The outer root sheath, however, is continuous with the surface epidermis at the top of the follicle.
The outermost layer of the hair is a cuticle like bark on a tree, more easily seen in cross sections (see below). The cuticle of the hair is so closely pressed against the hair shaft and the adjoining cuticle of the root sheath (see below) that the two are indistinguishable in your sections.
Inside the cuticle most hairs (not all) have two distinct regions: a cortex in which the dead keratinized cells are very densely packed, and a medulla, in which they aren't so densely packed. These will be easy to tell apart, especially in slide 35. The cortex looks like a rind on a melon, with the medulla as the "meat" inside. Some hairs have no medulla; these are the curly "wool hairs" as opposed to the long "guard hairs" seen in furbearing animals. The "glassy membrane" is the basement membrane of the follicle, separating it from the CT around it.

This hair has been cut in cross section, at a level of the follicle where the inner (IRS) and outer (ORS) root sheaths are both visible. The inner sheath has several layers, and in this field the translucent tricohyaline granules of Huxley's layer are easily visible. These granules are markedly similar to the keratohyaline granules of the epidermal keratinocytes: more proof that hair production is a similar process. In this picture the deeply pigmented cortex and less pigmented medulla of the hair are visible. This is from slide 35, on which most of the hairs are cut in cross section.
 This hair, also from slide 35, is cut in cross section above the point in the follicle where the inner root sheath terminates. Only the outer root sheath and the hair proper are visible. Notice the small sebaceous gland profile at lower right.
This hair, also from slide 35, is cut in cross section above the point in the follicle where the inner root sheath terminates. Only the outer root sheath and the hair proper are visible. Notice the small sebaceous gland profile at lower right.
Some hairs are hollow. Instead of a cellular medulla, there's an air space inside. This is pretty common in Arctic animals, most notably the polar bear. By having an air space inside the hair, its insulation value is greatly increased. This eons-old natural solution to the problem of keeping out the cold even has a human made equivalent: the commercial fiber "Holofil" used in sleeping bags and low cost insulated jackets. It's basically imitation polar bear hair, a thin acrylic filament with a hole through the middle. Imitation is indeed the sincerest form of flattery.
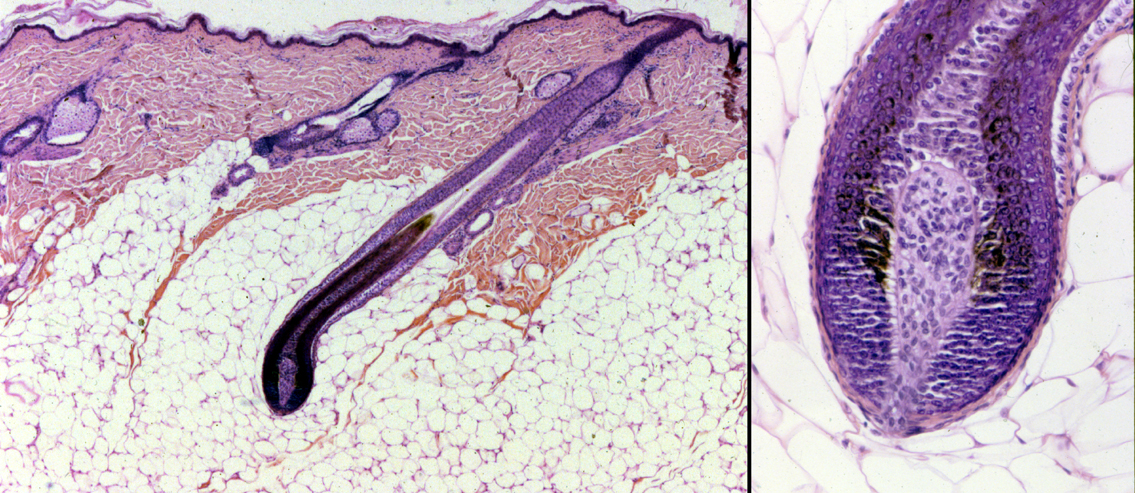
Hair Bulb and Dermal Papilla
At the bottom of the follicle the epithelium swells to form a hair bulb. This is the dilated end from which the growth of the hair and the inner root sheath occur. The hair bulb in an active hair follicle is usually large enough to enclose a bit of the dermis. This is the dermal papilla of the follicle, which plays an important role in the hair growth cycle.
The dermal papilla itself isn't the source of the hair, the active epithelium lying over it is. But the papilla is necessary to hair growth, in that its presence and close proximity to this germinal matrix induces it to proliferate by sending chemical signals. When the signal is received, the germinal matrix proliferates and produces the hair and the inner root sheath (see below). You can think of the germinal matrix as the homolog of the stratum basale of the epidermis, because it's here that everything is produced. Another similarity is that it contains melanocytes; just as in the epidermis, the melanocytes of the germinal matrix transfer melanin to hair to give it pigmentation.
The triggering of hair growth by proximity of the dermal papilla is a classic example of the phenomenon of induction, the response of one cell population to signals from a second population. You may notice that the bulbs and papillae in a given section of skin are not all the same size. You can expect to see smaller papillae in hairs which are being shed. The catagen phase is characterized by a shrinkage and withdrawal of the papilla away from the bulb. This removes the necessary induction and the hair stops growing.
Everything discussed so far has been epithelial in nature. But the large hair follicle is embedded deep in the nonepithelial dermis, and CT structures play an important role. The epithelium of the follicle has to rest on a basement membrane, and you will see this as a thickened amorphous structure right outside the outer root sheath. In this case the term applied to the basement membrane is the glassy membrane. It's surrounded in its turn by the less dense dermal sheath which is with the CT of the dermis.
Hair Growth
A hair in and of itself is dead material, but it's produced by living cells. Those cells are epidermal in nature and origin, and the hair, like the other adnexa of the skin, is epidermal in nature. Specialized keratinocytes at the base of the hair produce the hair shaft in a process that's very similar to the keratinization of the surface epidermis. The pigmentation in hair comes from melanocytes, just as the pigmentation of the skin does. The follicle's outer layers are continuous with the surface epidermis: a hair doesn't grow "through" the skin, it grows from the skin. It's part of the skin's complete structure.
Patterns of hair growth and degrees of hairiness vary among species, and even among individuals of the same species. Of course, like any genetically determined characteristic, hair can be manipulated by man for his own purposes through selective breeding. Many animals are raised specifically for their pelts: minks, chinchillas, ermines, nutria, and sheep, to name only a few.
Hair growth in animals is intimately related to seasonal cycles, and in most cases the cycle of hair growth and quiescence is cued by changing day length. Some animals change hair color or length in response to changes in day length (example, the varying hare) and most undergo a cycle of shedding and re-growth, a phenomenon with which every dog owner is familiar.
 The cycle of hair growth involves an active phase of production, anagen, during which the hair grows in length by addition of cells to the bottom (deep) end. At some point or other, influenced by a number of factors, this stage ends and is replaced by catagen, the stage of quiescence and cessation of new hair cell production.
The cycle of hair growth involves an active phase of production, anagen, during which the hair grows in length by addition of cells to the bottom (deep) end. At some point or other, influenced by a number of factors, this stage ends and is replaced by catagen, the stage of quiescence and cessation of new hair cell production.
The hair in catagen may (and usually does) detach itself from the underlying matrix which produced it, and be held in the follicle simply by friction. This is a club hair. Club hairs aren't anchored in so they fall out very easily. That's where the hairs embedded in the bristles of your hairbrush come from every morning, and where the hair on your bed comes from if you have a Siberian Husky in the house.
Eventually something wakens the follicle from its dormancy, and it enters into the transitional stage of telogen, during which it renews itself for activity, and makes the changeover back to the active anagen stage again.

Hair is affected by any number of things: nutrition, age, health status, season of the year, sex, and many other factors. Many of these are controlled by hormonal flux. The most familiar hormonal effect hair growth is the differences in hair growth in males and females. In humans "male pattern baldness" is due to the suppressive effect testosterone exerts on the follicles of the scalp. It is not normally seen in women, because their testosterone levels are so low. In extremely old women, whose sources of estrogen have completely atrophied, baldness of the male pattern type does occur. The drug minoxidil takes advantage of the responsiveness of hair follicles to hormones.
Testosterone has just the opposite effect on hair follicles of the face. In men it's the onset of puberty and the elevated levels of testosterone that stimulate the follicles of the face to make beard hairs, and beards are exclusively a masculine feature. Extremely old women can grow stubble like a man's when their estrogen dries up, because women do produce very small amounts of testosterone. When their source of estrogen is completely gone, there's enough to stimulate their facial hair follicles.
 Equivalent phenomena occur in animals. The "ruff" of fur around the neck is associated with some dog breeds in intact males, as is the large and impressive mane of the male lion. In gorillas, maturation of males leads to the development of silvery tips on the guard hairs, and a male that has reached a certain stage is referred to as a "silverback."
Equivalent phenomena occur in animals. The "ruff" of fur around the neck is associated with some dog breeds in intact males, as is the large and impressive mane of the male lion. In gorillas, maturation of males leads to the development of silvery tips on the guard hairs, and a male that has reached a certain stage is referred to as a "silverback."
Color change with season is another hormonally-controlled aspect of the hair cycle: in that instance it's the activity of melanocytes being modulated to make more or less pigment. The varying hare of the northern latitudes shuts its melanocytes off in the Winter, and grows white hair as camouflage in the snow; in Spring the melanocytes ramp up production again and the hare turns brown, to better blend in with the foliage on the forest floor.
Other things that influence hair growth include electromagnetic radiation (especially hard gamma rays and X-rays) and many drugs. Any agent, physical or chemical, which affects rapidly dividing cells will affect hair growth, usually by stopping it. Hair loss (alopecia) is a common side effect of radiation treatment and chemotherapy. Parasites, disease, and poor nutrition all affect hair growth. The condition of an animal's hair coat is usually a very good diagnostic clue to its internal medical problems.
Sinus Hairs
Sinus hairs are very large hairs modified to act as mechanical sensors. The "whiskers" or vibrissae on dogs and cats are good examples. Structurally they are identical to "normal" hairs except for size. Slide 1203 will give you an opportunity to examine the structure of sinus hairs. These are extremely large hairs, and they have large, blood filled sinuses surrounding their bases.
 A dog or cat has vibrissae on the sides, top, and bottom of his head. These serve the same function as curb feelers on a car, i.e., they alert the animal when he's close to an edge. This keeps the animal from getting his head stuck in a bucket or a hollow tree. Cats never get their heads stuck, but most dogs I've known have blithely ignored the signals, and managed to do so at least once in their lifetime.
A dog or cat has vibrissae on the sides, top, and bottom of his head. These serve the same function as curb feelers on a car, i.e., they alert the animal when he's close to an edge. This keeps the animal from getting his head stuck in a bucket or a hollow tree. Cats never get their heads stuck, but most dogs I've known have blithely ignored the signals, and managed to do so at least once in their lifetime.
Actually, all hairs have fine nerve fibers which invest the base of the follicle, and serve as tactile sensors. If you don't believe this, pull one out of your arm.