Rods and Cones
Role of the Rods and Cones
The rods and cones (strictly speaking, those portions that
are outside of the outer limiting "membrane" of the retina) are the actual sites
of transduction of light energy into neuronal signals. They are, in essence,
exceptionally specialized bipolar neurons, which have developed some structural
features to carry out this task.
The rod is the "model" transducer, and
will be given most attention here. The mechanism by which
light energy is converted to neuronal signals is exactly the same in both
rods and cones; the difference between the two types of receptor are in the
visual pigments involved.
Rods respond to very low levels of light at all wavelengths
of the visible spectrum by generating a signal. Consequently they are of greatest
importance under conditions in which lighting is dim and discriminating colors
is not a primary requirement. However, the cones are wavelength specific to
a degree, and are therefore responsible for color vision. Cones are also much
less sensitive than rods, and require higher levels of light to generate signals.
Thus they work best in daytime conditions.
Structure of the Rods and Cone
 Each is a fairly large cell. The rods and cones are "polarized"
in the sense that they have distinctly different architecture and function at
each end. The outermost portion of these cells is a highly modified cilium,
greatly expanded in size, and consisting of a stack of numerous light-sensitive
folds of membrane material. These stacked lamellae are the actual sight of transduction.
The names "rod" and "cone" reflect the general shape each type of light receptor
takes, and in good preparations for the light microscope, the shapes are easily
visible.
Each is a fairly large cell. The rods and cones are "polarized"
in the sense that they have distinctly different architecture and function at
each end. The outermost portion of these cells is a highly modified cilium,
greatly expanded in size, and consisting of a stack of numerous light-sensitive
folds of membrane material. These stacked lamellae are the actual sight of transduction.
The names "rod" and "cone" reflect the general shape each type of light receptor
takes, and in good preparations for the light microscope, the shapes are easily
visible.
What is shown at left is actually only part of the cells,
the outer segment of each type. This image, which is based on detailed
reconstructions from transmission electron micrographs, clearly shows the origins
of the names "rod" and "cone". The outer segments drawn here, are connected
by a narrow "waist" or constriction to the inner segment of each cell
type. Both outer and inner segments are physically isolated from the cell bodies
of the rod and cone cells by the outer limiting "membrane" and the actual cell
body is located in the outer nuclear layer.
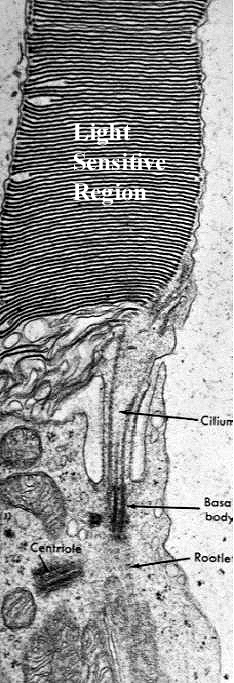 The
image at the left, a transmission electron micrograph, shows the "waist" between
the inner and outer rod segments. The light sensitive lamellae of the outer
segments are obvious, as is the narrow constriction. Note that in the constriction
itself, there is an actual modified cilium, complete with basal body and ciliary
rootlets. This remarkable structure is proof that the outer segment is really
a highly modified cilium, and that the membranous lamellae are extensive ramifications
of its plasma membrane. the modification of cilia to perform sensory functions
is found in other locations in mammals: the olfactory cells of the nose are
another example of a highly-specialized neuron with greatly-modified cilia devoted
to sense perception rather than movement of fluid.
The
image at the left, a transmission electron micrograph, shows the "waist" between
the inner and outer rod segments. The light sensitive lamellae of the outer
segments are obvious, as is the narrow constriction. Note that in the constriction
itself, there is an actual modified cilium, complete with basal body and ciliary
rootlets. This remarkable structure is proof that the outer segment is really
a highly modified cilium, and that the membranous lamellae are extensive ramifications
of its plasma membrane. the modification of cilia to perform sensory functions
is found in other locations in mammals: the olfactory cells of the nose are
another example of a highly-specialized neuron with greatly-modified cilia devoted
to sense perception rather than movement of fluid.
Vision is a vitally important sense in all vertebrates, but
some groups have better vision than others. Color vision is best in birds and
primates. Humans, whose senses of smell and hearing are less acute than those
of most quadrupeds, depend primarily on vision for orientation to the world
around them. Human vision is among the best in the animal kingdom, and the ability
of humans to detect subtle variations in colors is well documented. This makes
some evolutionary sense: in both groups the sense of smell is rather poor (many
birds are believed to have no sense of smell at all) and the necessity to identify
food sources and potential mates is crucial to survival. The bright colors of
birds serve as a visual means of species recognition to prevent unproductive
cross-specific matings. In primates, vision is used to identify edible plants,
and binocular vision confers the depth perception needed
for arboreal life.
There is a widely-held belief that domestic animals are "color-blind,"
but this has yet to be definitively proven. Such data can only be derived from
indirect experimentation using behavioral studies, but many such studies are
contradictory and inconclusive. It is certainly true that in some animals chemorecption
and hearing are more important sensory modalities than vision, but it is has
also been demonstrated that the color-sensitive cones are present in virtually
all animals to some degree. The anatomic structures for color vision are present.
Whether the signals these cells generate are interpreted as "color" in the same
sense that primates do is a matter of debate and certainly occurs at
the level of the central nervous system, not in the eye itself.
Click Back to return
Or Go To:
Main Page | Corneoscleral Tunic | Uveal Tunic | Retinal Tunic | Physiology of Vision | CNS Processing of Visual Signals