The Retinal Tunic
Regions of the Retinal Tunic
The term "retina" is almost always taken to mean the portion
of the eye's innermost tunic that's sensitive to light. And transduction
of light into nervous impulses is the chief function of the retina, without
doubt. In a sense, the retina is the eye: everything else is there simply
to ensure that the sensitive part of the retina is properly maintained and that
an image is brought to it in focus.
Insensitive Retina and the Ora Serrata
 But
in addition to this function, the retina has an insensitive region, a
portion whose elements do not respond to light rays and have other functions
in the normal eye. The line of demarcation between the sensitive and insensitive
portions of the retinal tunic is the ora serrata, a radial zone about
three-quarters of the way towards the anterior aspect of the globe. It's indicated
in the image at left by the arrow. The insensitive region,
about 25% of the retinal tunic, is devoted to participating in the production
of aqueous humor. The remaining 75% is dedicated to the eye's job of transducing
electromagnetic energy into nervous signals, and it's described in more detail
below.
But
in addition to this function, the retina has an insensitive region, a
portion whose elements do not respond to light rays and have other functions
in the normal eye. The line of demarcation between the sensitive and insensitive
portions of the retinal tunic is the ora serrata, a radial zone about
three-quarters of the way towards the anterior aspect of the globe. It's indicated
in the image at left by the arrow. The insensitive region,
about 25% of the retinal tunic, is devoted to participating in the production
of aqueous humor. The remaining 75% is dedicated to the eye's job of transducing
electromagnetic energy into nervous signals, and it's described in more detail
below.
Sensitive Retina
The business of the eye takes place in the sensitive region,
covering three-quarters of the innermost tunic. It is here that the image is
formed, in exactly the same way that a camera's lenses cast an image onto a
sheet of film. But the retina does much more than any film, because the initial
signal processing and interpretation takes place here. Consequently it is a
very complex structure.
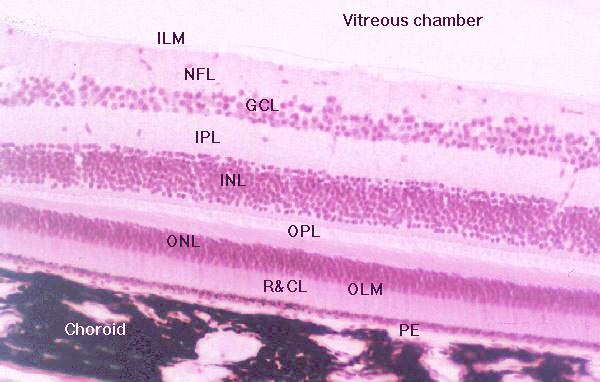
Layers of the Retina
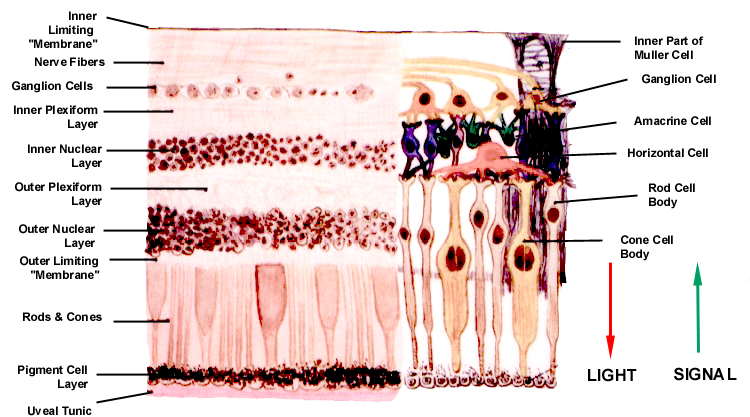
The vertebrate retina has nine layers whose origins
are from the inner wall of the primitive optic cup. A tenth layer, the pigment
cell layer, lies just outside these nine, and is so closely associated with
it in structure and function that it can be considered a part of the retina.
In vertebrates the light-sensitive elements are in the outermost
of the nine layers, and rays of light forming the image must pass through the
other eight layers before the nervous signal can be generated. In other words,
the "raw" image is actually formed at the back (the outer part) and the "processed"
signal that results is routed towards to the inner portion of the globe inward
by converging axons from a series of integrator cells located in the inner portions
of the retinal tunic.
Perhaps the most logical way to understand the progression
of events in the transduction of vision is to consider the layers of the retina
in sequential order, beginning at the outermost, and following the signal back
through the tunic to the point of exit via the optic nerve.
Pigment Epithelium Layer (PE)
This layer has vital functions in increasing acuity of vision and in
ensuring that the light sensitive portions of the rods and cones are maintained
in working order. It is a single layer of cuboidal cells.
These cells are impregnated with melanin, but they also contain
lipofuscin as a product of their metabolic activity. The light-sensitive parts
of the retina are continually "turned over" to maintain optimum function. The
pigment layer cells phagocytose the ends of the rods and cones as they are renewed:
hence the accumulation of lipofuscin. The lipofuscin and melanin complement
the pigmentation of the choroid portion of the uveal tunic.
Together these two layers increase the contrast of the visual image by absorbing
light that would otherwise be reflected back inwards towards the rods and cones.
The pigment epithelium forms part of the blood-retina barrier and in some "lower" vertebrates may be able to regenerate the retina proper.
Another and even more important function of the pigment epithelium
is the storage and synthesis of trans-retinal, or vitamin A.
This vitamin is reversibly convertible into trans-retinal, and
serves as a raw material for the cyclic formation and breakdown
of rhodopsin, the visual pigment.
Layer Rods and Cones (R&CL)
The outermost layer of the retina "proper" (i.e., the
nine that are formed from the inner part of the optic cup) is the bacillary
layer or the layer of rods and cones. These are
the actual light-sensitive elements. Rods and cones are a variant form of neuronal
cell, whose light sensitive portions represent one of the many examples of the
modification of cilia for sensory purposes.
The Outer Limiting "Membrane" (OLM)
The so-called outer limiting "membrane" isn't
really a membrane at all. Instead, it's the site of numerous occluding junctions.
These seal off the "disposable" parts of the rods and cones, the actual light-sensitive
portions of these cells, from the rest. The junctions are between the plasma
membranes of the rod segments and a glial element, the Müller
cell.
This outer limiting layer has the spurious appearance
of a "membrane" in light microscopic preparations, because there are so many
junctions, all closely packed together. The electron microscope reveals their
true nature. They serve to isolate the inner layers of the retina from potentially
harmful material in the blood circulation, forming a blood-retina barrier.
The Outer Nuclear Layer (ONL)
This layer is the location for the nuclei and
cell bodies of the rod and cone cells (whose sensitive elements project outwards
as the bacillary layer). Collectively the nuclei of these cells constitute
the outer nuclear layer.
The Outer Plexiform Layer (OPL)
Inwards of the outer nuclear layer is a relatively
clear zone. This is the site of numerous synapses between the rod and cone
cells and the processes of various integrator neurons, the outer plexiform
layer.
Once the initial neural signal is generated in
the rods and cones, it must be passed to other neural elements for processing
and further handling. The dendrites of these cells and the synaptic
portions of the rod and cone cells constitute the outer plexiform layer,
a region of synapses.
The integrating elements are the horizontal, bipolar,
and amacrine cells. The dendrites of the bipolar cells, in particular,
comprise most of the outer plexiform layer.
The Inner Nuclear Layer (INL)
The cell bodies and nuclei of the integrator
neurons, particularly the bipolar and horizontal cells, are located in the
inner nuclear layer, next inwards from the outer plexiform layer.
The Inner Plexiform Layer (IPL)
The inner plexiform layer, like the outer one,
is a region of synapses. Here the bipolar cell processes synapse with the
dendritic processes of ganglion cells, the final neuronal element of
the eye itself.
The Ganglion Cell Layer (GCL)
While the ganglion cells have synapses with
bipolar cells in the inner plexiform layer, their cell bodies are located
in the next retinal layer, the ganglion cell layer, which has far fewer nuclei
than the inner or outer nuclear layers. Like other neurons, these ganglion
cells have axons, which carry the generated signal. These axons are bundled
into tracts that run radially, forming the next layer of the retina.
The Nerve Fiber Layer (NFL)
The axonal fibers from the ganglion cells, bundled
together, run radially around the inner surface of the retina, converging
at the site of origin of the optic nerve. At that point the nerve penetrates
the retina and the sclera. At this point there are no light sensitive elements;
hence this is the "blind spot" found in all vertebrate
eyes.
The Inner Limiting "Membrane" (ILM)
The innermost layer, again, is not really a
membrane, but is instead a place where the foot processes of the Müller cells come together.
The body of the elongated Müller cell runs from one side of the retina to the other, and "fills in the spaces," so to speak, between the other elements.
Click Back to return
Or Go To:
Main Page | Corneoscleral Tunic | Uveal Tunic | Retinal Tunic | Physiology of Vision | CNS Processing of Visual Signals